HOOFDSTUK 9:
HERSENEN EN BEWUSTZIJN: VAN PNEUMA TOT GRIJZE MASSA
Wim van de Grind & Gert-Jan Lokhorst
1999
W. N. A. van de Grind & G.J.C. Lokhorst.
Hersenen en bewustzijn: van pneuma tot grijze massa. In Frank
Wijnen & Frans Verstraten, eds., Het brein te kijk:
verkenning van de cognitieve neurowetenschappen, pp. 217-246.
Swets en Zeitlinger, Lisse, 2001. ISBN 90-265-1676-2. Tweede druk:
2002. Derde druk: 2004.
Those who see any difference between soul and body have
neither
Oscar Wilde, Chameleon, 1894
We beginnen in 8.2 met een korte inleiding in de filosofische
aspecten van het hersenen-bewustzijn probleem en het probleem van
de vrije wil. Daarna gaan we in 8.3 in op de grenzen en
mogelijkheden van de studie van bewustzijn als een biologisch
verschijnsel en als een aspect van de werking van de hersenen.
Vervolgens bespreken we in 8.4 de eerste experimenten waarin werd
getracht een brug te slaan tussen activiteiten in het zenuwstelsel
en de bewuste ervaring en vrije wil. Dit bleek qua houdbaarheid van
de conclusies geen onverdeeld succes en neurowetenschappers hebben
erdoor geleerd wat voorzichtiger met de interpretaties van hun
bevindingen om te springen. Grote denkrichtingen en vergaande
conclusies worden tegenwoordig meestal vermeden. Dat heeft geleid
tot de meer pragmatische invalshoek die in het huidige
hersenonderzoek de boventoon voert. Dit pragmatisme blijkt het
bewustzijnsonderzoek voor het eerst in de geschiedenis op
wetenschappelijk acceptabele wijze binnen het domein van de
natuurwetenschap te brengen. Vooral onderzoek aan de visuele
perceptie lijkt toegang te verschaffen tot het `neuronaal correlaat
van bewustzijn'. In 8.5 geven we daarvan een overzicht om dan in
8.6 met enkele conclusies en een vooruitblik naar de toekomst te
eindigen.
Marietje zit achter haar computer en ziet een aantal letters op
het scherm staan. De fysica en de neurofysiologie geven een
tamelijk duidelijk beeld van wat er hierbij gebeurt: het
elektronenkanon in de monitor vuurt elektronen op het scherm af, er
komen fotonen vrij die naar haar ogen vliegen, er komen bepaalde
elektrochemische processen in haar netvlies op gang, er gaan
signalen naar de hersenen, en uiteindelijk vinden er zekere
veranderingen van de activiteiten in bepaalde gebieden van de
hersenen plaats. Maar de fysica en de neurofysiologie lijken het
interessantste aspect van dit gebeuren buiten beschouwing te laten,
namelijk het feit dat deze veranderingen van hersenactiviteit
gepaard gaan met bewuste gewaarwordingen in Marietjes geest.
Zouden die gewaarwordingen niet net zo goed niet kunnen
plaatsvinden? Het elektronenkanon is zich er per slot van rekening
ook niet van bewust dat het elektronen afvuurt. En waarom leidt het
fotonenbombardement van de retina eigenlijk tot visuele
gewaarwordingen? Waarom ziet Marietje een A op het scherm
als een gebied met een bepaalde kleur en vorm, in
plaats van dat ze hem ruikt als iets waar een bepaalde
karakteristieke geur vanaf komt? Ziedaar één
aspect van het hersenen-bewustzijn probleem, het probleem dat ons
in dit hoofdstuk zal bezighouden.
Het hersenen-bewustzijn probleem heeft nog een ander aspect.
Stel dat de letters die Marietje op het scherm ziet haar niet
bevallen. Zij kan dan besluiten om bepaalde veranderingen aan te
brengen. En inderdaad, even later voert zij de gewenste
veranderingen via het toetsenbord in. Hoe kan dit? Ons brein vormt
een ingewikkeld systeem dat in nauwe samenhang met de omgeving
voortdurend actief is. Op die activiteit lijkt het `ik' dat de
eigenaar van het brein denkt te zijn geen invloed uit te kunnen
oefenen. En zelfs als `ik' al invloed zou kunnen uitoefenen, dan
zou `ik' nog niet eens weten hoe ervoor te zorgen dat de activiteit
zodanig verandert dat de vingers de gewenste beweging uitvoeren.
Hoe kan onze wil dan invloed hebben? Heeft ze die eigenlijk
wel?
Het moderne hersenen-bewustzijn probleem dateert uit de
zeventiende eeuw en is vooral te danken aan Descartes. Descartes
maakte een scherp onderscheid tussen de materie aan de ene kant en
de ziel aan de andere kant. Hij beschouwde de ruimtelijke
uitgebreidheid als het essentiële kenmerk van de materie en
het denken als het essentiële kenmerk van de ziel; hij meende
dat de materie niet kan denken en dat de ziel geen ruimtelijke
afmetingen heeft. Hij meende overigens niet dat de ziel
alléén maar kan denken: hij dacht dat ze ook kan
waarnemen, willen, gevoelens kan ondergaan, zich gebeurtenissen kan
herinneren, en zo voort. Kortom, alles wat we tegenwoordig als
`bewuste psychische (of mentale) verschijnselen' zouden willen
bestempelen kwam volgens Descartes toe aan de onstoffelijke ziel.
Het lichaam daarentegen was volgens Descartes niets meer dan een
machine, een automaat die op een mechanische manier zijn eigen gang
gaat in overeenstemming met de wetten van de natuur. Deze opvatting
paste goed in de `mechanisering van het wereldbeeld' die destijds
in volle gang was (Dijksterhuis, 1950).
Descartes maakte een dermate scherpe scheiding tussen ziel en
lichaam dat het moeilijk was om zich te voor stellen hoe de twee
verband met elkaar kunnen houden. Enerzijds was het niet duidelijk
hoe gebeurtenissen in het lichaam tot psychische verschijnselen
zouden kunnen leiden; anderzijds was het moeilijk voorstelbaar hoe
psychische verschijnselen van invloed zouden kunnen zijn op de
cerebrale machinerie.
Vóór de zeventiende eeuw leefden deze problemen
minder sterk omdat men zowel een ander beeld van de materie als een
ander beeld van psychische verschijnselen had. Aan de ene kant zag
men de natuur nog niet als een machine die net zo mechanisch
functioneert als een uurwerk. In de natuurkunde van Aristoteles
bijvoorbeeld hadden materiële voorwerpen als het ware hun
eigen verlangens en antipathieën; ze streefden er bijvoorbeeld
naar om hun `natuurlijke plaats' op te zoeken. De materie was dus
lang niet zo `zielloos' als in de opvatting van Descartes. Aan de
andere kant werden psychische verschijnselen ook niet zonder meer
als onlichamelijk beschouwd. Plato, Aristoteles en de middeleeuwse
scholastieke filosofen beschouwden alleen het verstand als
onstoffelijk en onsterfelijk; de andere hogere psychische
vermogens, zoals waarnemen en herinneren, werden in nauw verband
gebracht met de materie. Vanaf ongeveer de derde eeuw voor Christus
(toen men in Alexandrië voor het eerst goed anatomisch
onderzoek begon te verrichten) meende men over het algemeen dat ze
verbonden waren met een heel fijn of etherisch materiaal, in het
Grieks pneuma psuchikon en in het Latijn spiritus
animalis genaamd. Deze gasachtige substantie zou door de
hersenholten (ventrikels) en de zenuwen (die men zich als hol
voorstelde) waaien en verantwoordelijk zijn voor de waarneming, het
denken en de motoriek. Deze opvatting beheerste het denken tot in
de zeventiende eeuw (Verbeke, 1945; Clarke en O'Malley, 1968;
Clarke en Dewhurst, 1973; Finger, 1994). De `animale geesten'
speelden ook een rol in de theorie van Descartes. Net zoals zijn
voorgangers dacht hij dat de hersenholten een `wind of vlam'
bevatten die bemiddelde tussen de hersenen en de rest van het
lichaam; maar hij benadrukte dat de `esprits animaux' uit gewone,
gedachteloze, aan de wetten van de mechanica onderworpen materie
bestonden.
De fysiologische details van Descartes' theorie doen
tegenwoordig vreemd aan. Hij dacht dat de ziel voornamelijk
verbonden was met de pijnappelklier, een klein orgaantje in het
midden van de hersenen. (Dit idee was overigens veel origineler dan
gewoonlijk wordt beweerd: zie Lokhorst & Kaitaro, te
verschijnen.) De zenuwen zouden holle buisjes zijn waarin draden
lopen die prikkels vanuit de zintuigen naar de pijnappelklier
kunnen overbrengen; de prikkelingen van de pijnappelklier zouden
vervolgens kunnen worden geregistreerd door de onstoffelijke ziel,
en zo zouden bewuste waarnemingen ontstaan. Soms zou de ziel de
pijnappelklier een bepaalde kant opduwen, waardoor de esprits
animaux van richting zouden veranderen en bepaalde spieren
opgeblazen zouden kunnen worden: op die manier zouden bewuste
beslissingen van de ziel tot bewegingen van het lichaam kunnen
leiden.
Maar al zijn de fysiologische details van Descartes' theorie
vreemd en verouderd, de filosofische strekking van zijn theorie is
nog steeds van belang en heeft een grote invloed tot op de dag van
vandaag. Er zijn nog steeds geleerden die menen dat de mens uit
twee componenten bestaat, een materieel lichaam en een
onstoffelijke ziel, die in een causale interactie staan op de door
Descartes beschreven wijze. En er zijn al meer dan drie eeuwen lang
geleerden die hun best doen om deze theorie onderuit te halen. Ja,
Descartes' interactionistisch dualisme heeft in feite al meer dan
drie eeuwen lang als dé Kop van Jut gefungeerd in de
filosofie van de geest.
Al in de zeventiende eeuw kreeg Descartes' theorie meer kritiek
dan bijval. Er valt onder meer het volgende tegen in te
brengen.
Ten eerste verklaart de theorie helemaal niets. Het is
immers niet minder onbegrijpelijk hoe een immateriële ziel zou
kunnen waarnemen, denken, willen, enz., dan hoe een materieel ding
zoals de hersenen dat zou kunnen doen.
Ten tweede creëert de theorie alleen maar nieuwe
problemen: we zouden immers graag willen weten hoe de
immateriële ziel met het materiële lichaam in
wisselwerking treedt, maar hier heeft geen enkele dualist ooit iets
zinnigs over gezegd.
Ten derde is er geen enkel geprivilegieerd punt in de
hersenen waar de ziel speciaal mee verbonden is. De pijnappelklier
is vaak versteend (zoals men al in de zeventiende eeuw ontdekte),
het corpus callosum (de favoriete plaats van de ziel in de
achttiende eeuw, zoals bijvoorbeeld blijkt uit het artikel over de
ziel in de Encyclopédie van Diderot) kan zonder
fatale gevolgen worden doorgesneden (zoals Zinn in de achttiende
eeuw aantoonde), er is geen enkele zenuwcel waarvan het uitvallen
in zijn eentje tot uitval van alle mentale activiteit leidt, enz.
enz.
Ten vierde is de ziel niet één en
ondeelbaar: allerlei specifieke vermogens (bijvoorbeeld
waarneming van beweging, niet van plaats) kunnen uitvallen terwijl
de rest van de psyche intact blijft. We kunnen de bewustzijnsstroom
zelfs doorsnijden (verdubbelen) door de hersenen door te snijden,
zoals het split-brain syndroom (dat mensen vertonen bij wie het
corpus callosum is doorgesneden) laat zien (Lokhorst,
1996).
Ten vijfde staat de theorie op gespannen voet met de
natuurkunde. De natuurwetten gelden ook in de hersenen; er is
geen enkele reden om aan te nemen dat er een niet-fysische
instantie is die ervoor zorgt dat ze in de hersenen niet gelden
doordat zij af en toe een vinger in het raderwerk steekt.
Tegelijkertijd lijken we ook geen speciaal fysisch principe
(een psychisch veld of iets dergelijks) nodig te hebben om de
werking van de zenuwen te verklaren. Er is dus gewoonweg geen
plaats voor een speciaal psychisch beginsel van niet-fysische dan
wel fysische aard.
In de loop van de tijd zijn de meeste filosofen tot de
overtuiging gekomen dat de hersenen het orgaan zijn waarmee we
waarnemen, denken, leren, vergeten en bewegen. Mentale processen
(zoals ergens over nadenken) zijn hersenprocessen, mentale
gebeurtenissen (zoals schrikken van een geluid) zijn gebeurtenissen
in de hersenen, en mentale toestanden (zoals geagiteerd zijn) zijn
hersentoestanden.
Dit wil niet zeggen dat je mentale termen kunt
definiëren in hersentermen, dat je mentale taal kunt
vertalen in hersentaal, of dat je de psychologie kunt
reduceren tot neurobiologie: ook al is ieder psychologisch
fenomeen een hersenfenomeen, dat wil niet zeggen dat er een
wetmatige samenhang bestaat tussen beide domeinen. Denk aan
picknicks: iedere picknick bestaat uit atomen, maar je kunt
picknicks niet in fysische termen definiëren, je kunt
beschrijvingen van picknicks niet vertalen in de taal van de
fysica, en je kunt de picknick-kunde of -kunst niet
reduceren tot natuurkunde. Met een typisch mentaal fenomeen
als `denken aan het zinken van de Titanic' is het wellicht niet
anders gesteld. Iedere gedachte aan het zinken van de Titanic is
wellicht een fysisch fenomeen, maar dat impliceert niet dat we
`denken aan het zinken van de Titanic' ooit in de taal van de
neurofysiologie zullen kunnen definiëren.
De sterkste niet-controversiële algemene uitspraak die we
kunnen doen is wellicht deze: geen psychisch verschil zonder
fysisch verschil. Het fysische bepaalt het psychische.
Filosofen spreken in dit geval van superveniëntie: ze
zeggen dat het psychische supervenieert op het fysische. De
stelling dat het psychische supervenieert op het fysische sluit
natuurlijk niet principieel uit dat andere wezens, wellicht gemaakt
van heel andere materialen dan wij, bijvoorbeeld silicium,
positronen (denk aan de breinen van de robots van Asimov) of
pneuma (de katholieke kerk leert nog steeds dat engelen uit
deze fijne materie bestaan), precies dezelfde waarnemingen,
gevoelens, gedachten en herinneringen zouden kunnen hebben als
wij.
Is het geest-lichaam probleem hiermee nu opgelost? Dat nog niet:
we weten immers nog steeds niet hoe elektrochemische activiteit in
de hersenen tot bewuste ervaringen leidt. Waarom verschijnt de
buitenwereld aan mij zoals ze dat doet, en waarom heb ik eigenlijk
ervaringen? Zou mijn lichaam niet op precies dezelfde wijze kunnen
functioneren als het nu doet zonder dat er sprake is van een
innerlijke bewustzijnsstroom? Dit zijn vragen die niet zomaar
beantwoord kunnen worden. Toch geeft het bovenstaande de richting
aan waarin filosofen het zoeken. Ze beseffen daarbij dat ze hun
beperkingen hebben. De filosofie kan het niet in haar eentje
redden. Als we inzicht willen krijgen in verschijnselen als
waarnemen, denken, leren en vergeten, hebben we de empirische
wetenschappen nodig. Daar zullen we onze aandacht in het
onderstaande dan ook op richten.
Voor een goede algemene introductie in de filosofie van de geest
verwijzen we naar Kim (1996).
Welke algemene visie houden empirische hersenonderzoekers er
grosso modo op na? Het ziet er naar uit dat de meesten zich kunnen
vinden in het beeld dat John Searle (1984, 1992, 1998) schetst.
Searle is een filosoof die voortdurend met beide benen op de grond
wil blijven staan, een houding die wel de `common sense' aanpak
wordt genoemd. Natuurlijk zijn niet alle neurowetenschappers het
daarmee eens: zelfs het Cartesiaanse dualisme is nog niet geheel en
al uitgestorven in de kringen van hersenonderzoekers (zie Lokhorst
1986). We beschrijven echter in het volgende de meningen die het
meest in de biologische en neurowetenschappelijke literatuur lijken
voor te komen en daar sluit de aanpak van Searle uitstekend bij
aan.
John Searle (1984, 1992, 1998) heeft o.a. getracht argumenten
aan te voeren tegen het idee dat het zenuwstelsel een `computer'
zou zijn, waarop we in 8.3.5 kort ingaan Hij heeft uitgebreid
beargumenteerd dat bewustzijn en andere mentale verschijnselen
hoger-niveau kenmerken van de hersenen zijn die door lagere niveau
neuronale processen worden veroorzaakt (9.3.6). De moeilijke vraag
is dan hoe neurobiologische processen in het brein precies
bewustzijn veroorzaken. In een recente publikatie (Searle, 1998)
stelt hij dat de neurowetenschappen toe zijn aan de aanpak van dit
probleem, maar dat er nog een aantal filosofische misverstanden
moet worden ontzenuwd die de vooruitgang belemmeren. Die
misverstanden volgens Searle bespreken we in 8.3.2 t/m 8.3.4 in de
context waarin ze het meest optreden. In de bespreking richten we
ons vooral op de groeiende consensus in neurowetenschappelijke
kring over de methoden en uitgangspunten waarop men de
experimentele studie van bewustzijn en vrije wil het beste kan
baseren.
Een eerste misverstand is volgens Searle (1998) dat bewustzijn
niet bestudeerd zou kunnen worden omdat we er geen goede definitie
van hebben. Het is goed mogelijk het soort van globale definitie te
geven die altijd voldoende is geweest om wetenschappelijk onderzoek
te starten. Oorspronkelijk definieerde men water ook gewoon als de
kleurloze en smaakloze vloeistof die tijdens regen uit de hemel
valt. Aan het eind van het onderzoek komt dan een analytische
definitie zoals water is H2O. Zo zou men
bewustzijn initieel kunnen definiëren als de toestand waarin
we verkeren na het wakker worden en voor het weer in slaap vallen
of in coma raken of sterven. We weten daarmee welke toestand
bestudeerd moet worden. Daarmee is een voldoende helder beginpunt
voor wetenschappelijk onderzoek van die toestand gevonden, aldus
Searle.
Laten we proberen dit iets verder uit te werken, want wakker
zijn is dan wel een voorwaarde voor bewust zijn, maar er zijn
gradaties van wakker zijn en van slapen of in coma zijn. Van de
Grind (1997) noemt deze hele dimensie het WC-bewustzijn
(Wakker-Coma) en betoogt dat de toestand op deze glijdende schaal
te maken heeft met de algehele activatietoestand van het brein. We
beschouwen dit als een relatief oninteressant aspect van het
bewustzijn, iets dat zeker met succes natuurwetenschappelijk kan
worden bestudeerd. We weten gewoon uit ervaring dat vertebraten
zonder bewustzijn niet eten of reproduceren, niet migreren en hun
territorium niet verdedigen (Baars, 1998). Coma en diepe slaap
geven een brein te zien met heel andere activiteitspatronen dan een
wakker (dus WC-bewust) brein. Bewustzijn in deze zin
(WC-bewustzijn) is een keihard neurowetenschappelijk gegeven, geen
subtiel verschijnsel. Het kan op de operatietafel prima gehanteerd
worden. Laten we het hier dus als een gegeven aannemen en kijken of
we een stap verder kunnen gaan, te beginnen bij bewuste
handelingen.
Er is in elk geval ook een groot verschil tussen automatisch
handelen (buiten de bewuste controle om) en bewust handelen, zowel
qua gedragskenmerken als qua hersenactiviteit. We zouden bewust
handelen kunnen definiëren als dat handelen waarvan we weten
dat we het doen en/of waarover we expliciet kunnen rapporteren. Nu
treedt hier een probleem op, omdat we onszelf ook kunnen waarnemen.
Het is dus denkbaar, dat we kunnen rapporteren dat we iets doen,
zonder dat we ons van het handelen op zich bewust zijn, doordat we
wel bewust (maar dus achteraf) waarnemen wat we aan het doen zijn.
Dit zou betekenen dat ons handelen volledig automatisch gebeurt,
dat we geen vrije wil hebben, geen bewuste beslissingen nemen die
voorafgaan aan het handelen en er de oorzaak van zijn. Velen menen
dat zo'n model houdbaar is en we zullen in 8.4 een goed voorbeeld
zien van dit ideeëngoed. Hierbij wordt `vrije wil' als illusie
van bewuste zelfwaarneming beschouwd. In 8.4 zullen we zien dat de
evidentie voor dit idee niet zo sterk is als velen menen of hopen.
Maar, zelfs als er geen bewuste handelingen zouden bestaan (geen
vrije wil) moet men toch bewuste (zelf-)waarneming aannemen om de
illusie van vrije wil te verklaren. Kunnen we dan een definitie van
bewuste waarneming geven die uitgaat boven de vaststelling dat de
waarnemer `wakker' moet zijn? We stellen in de geest van Searle de
volgende werkdefinitie voor:
Bewustzijn is een toestand van wakker zijn waarbij men onder
andere bewuste ervaringen (gevoelens, gedachten, waarnemingen) kan
hebben. Ervaringen zijn bewust als men ze eigener beweging of op
verzoek (dus niet reflexmatig of als automatisme) en
reproduceerbaar kan rapporteren, dus waarneembaar kan maken voor
anderen.
Dit is voldoende als startpunt en de definitie is geen wettekst
of bijbeltekst, die men letterlijk moet gaan nemen of waar men
allerlei verborgen interpretaties bij moet gaan verzinnen. De term
`onder andere' geeft aan dat we ons een uitbreiding kunnen
voorstellen die over `bewust handelen' in plaats van bewust
waarnemen of denken gaat. Dus handelingen zijn bewust als men ze in
principe kan rapporteren vóór ze waarneembaar zijn.
Als men kan rapporteren hoe de beslissing tot bewust handelen tot
stand is gekomen vóór men de handeling start is er
sprake van vrije wil. Met `rapporteren' bedoelen we natuurlijk niet
noodzakelijkerwijs het geven van een gesproken of gedrukt verslag
(al mag dat ook). Stel dat we een poefpersoon zouden vragen of een
proefdier zouden trainen de linkerarm of voorpoot op te tillen
zodra ze iets groens zien. Als ze dat dan inderdaad reproduceerbaar
doen als er iets wat wij groen noemen in principe door hen gezien
kan worden, kunnen we stellen dat ze een groenervaring hebben, een
bewust groenpercept. Ze kunnen het percept op verzoek of eigener
beweging en op reproduceerbare wijze aan de omstandigheden
gekoppeld rapporteren.
Natuurlijk moeten we dan nog uitsluiten dat het reflexmatig
gedrag is, geconditioneerd gedrag, een automatisme, maar dat is
meestal niet zo moeilijk. Immers dan zou het gedrag door moeten
gaan als het geen zin heeft in de context van de rapportage. Als
proefpersoon of proefdier een beloning van de proefleider krijgt
voor het signaleren van de groenervaring en het signaleren houdt op
als de proefleider weggaat of als de beloning niet langer op prijs
wordt gesteld of is opgebruikt, is er geen sprake van
geconditioneerd of reflexmatig gedrag. Het is weinig zinvol om
tekortkomingen in onze werkdefinitie te gaan zoeken, die zijn er
zeker. Het is direct duidelijk dat er vele grensgevallen denkbaar
zijn van ervaringen die de een wel en de ander niet bewust wil
noemen. Ook éénmalige ervaringen vallen er niet
onder. In de neurowetenschap is het reproduceerbaar eigener
beweging kunnen rapporteren als kern van het begrip bewuste
perceptie genomen en daarop zullen wij ons verder concentreren.
Daarbij laat men dus subtiliteiten buiten beschouwing. Het is zeker
interessant om je af te vragen of de genoemde groenervaring bij
verschillende individuen of diersoorten wel vergelijkbaar is. Kun
je ervaringen eigenlijk wel zinvol vergelijken? Komen vergelijkbare
ervaringen wel met vergelijkbare activatie van vergelijkbare
neuronale schakelingen overeen? Zo kan men veel interessante
vraagstukken formuleren. Echter de neurowetenschap richt zich
voorlopig vooral op dat wat experimenteel direct hanteerbaar is. De
subtiliteiten komen dan later wel.
Het is een misverstand te menen dat bewustzijn niet
experimenteel bestudeerd kan worden omdat het niet observeerbaar
zou zijn. Dat bewustzijn niet direct observeerbaar zou zijn is
zeker geen probleem, want dat geldt ook voor zwarte gaten, de Big
Bang en vele andere goed bestudeerde verschijnselen. Als de
consequenties observeerbaar zijn is dat voldoende en voor
bewustzijn, bewuste ervaringen van pijn, kleur, verliefdheid e.d.
zijn de consequenties observeerbaar of kunnen observeerbaar worden
gemaakt. Searle noemt verder het misverstand dat wetenschap zich
alleen met objectieve dingen bezighoudt, terwijl bewustzijn
subjectief is. Objectief betekent in de praktische wetenschap
echter alleen dat we kunnen vaststellen of de uitspraken waar of
onwaar zijn ongeacht onze voorkeuren, vooroordelen of instelling.
Iedere waarnemer met de juiste gereedschappen moet de waarneming
kunnen herhalen. Het is daarbij dus irrelevant dat we niet weten
hoe een bewuste ervaring bij het onderzochte organisme aanvoelt
(ontologisch subjectief is). We kunnen dat wat met bewuste
ervaringen correleert bemeten en daarover falsifieerbare
theorieën opstellen. Datzelfde geldt voor de zwaartekracht of
welk ander fysisch fenomeen dan ook. We kunnen er niet achter komen
wat het `is' in ontologische zin, maar we kunnen er aan meten en
mee werken. Dat is voor wetenschappelijk onderzoek voldoende. De
verschijnselen zijn hanteerbaar en worden door een theorie
beschreven en voorspeld. Daar gaat het om in de wetenschap.
Evenzo is het een misverstand te menen dat we nooit zouden
kunnen begrijpen hoe iets subjectiefs, zoals bewuste ervaringen,
voort zou kunnen komen uit iets objectiefs als hersenactiviteit.
Dat vormt weliswaar een interessante puzzel vergelijkbaar met de
aard van het leven of de aard van de zwaartekracht, maar is voor
wetenschappelijk onderzoek geen belemmering. We weten eenvoudigweg
dat het zo is, er is niets anders dat in aanmerking komt als
veroorzaker van bewuste ervaringen dan hersenactiviteit. Dat
uitgangspunt is voldoende om nader te kunnen analyseren welke
hersenactiviteit de veroorzaker is, kortom om neurowetenschappelijk
onderzoek aan het verschijnsel te doen.
Men moet dan ook volgens Searle de qualia (de waargenomen
kwaliteiten van de dingen, bijvoorbeeld roodheid bij het zien van
iets roods) niet scheiden van bewustzijn, omdat het probleem van de
qualia te moeilijk zou zijn. Zonder qualia is er geen
bewustzijnsprobleem stelt Searle, want bewustzijnstoestanden zijn
per definitie van de aard van qualia (=kwalitatieve toestanden). De
vraag hoe iets subjectiefs als de qualia voort kan komen uit iets
objectiefs als de hersenen is de kern van ons vraagstuk en daartoe
moeten we eerst vaststellen hoe ze samenhangen. Welke
hersenactiviteit correleert met welke qualia? Zo'n vraag kan
experimenteel worden aangepakt.
Het volgende misverstand is volgens Searle dat bewustzijn een
epifenomeen is, een gevolg van de werking van het zenuwstelsel dat
zelf geen causale invloed uitoefent op de voortgang van die
activiteit. Beschouw, zo stelt Searle voor, de zuigers van een
automotor. Die zijn van vaste harde stof en dat is een gevolg van
de microstructuur van de materie. Toch is het geen epifenomeen,
want dankzij die macroscopische eigenschap van hardheid kan de
zuiger gebruikt worden in de motor en dus weer gevolgen hebben voor
de machine, auto en gebruikers. Voor de motor zijn de
macroscopische eigenschappen relevant en de oorzaak van allerlei
interacties. Dat ze veroorzaakt worden door microscopische
eigenschappen is daarbij irrelevant, want die worden weer
veroorzaakt door submicroscopische en subsubmicroscopische
processen, maar redenerend in gluonen (elementaire
materiesplinters) kun je geen automotor ontwerpen. Zo ook hebben
bewuste processen effecten, causale invloeden, die we niet aan
neuronen en aan eiwitten in neuronmembranen gaan toeschrijven, ook
al komen ze daar indirect uit voort. Wie zegt dat je de causale
ketens in alle richtingen tot in het oneindige terug moet
vervolgen? Dan zijn de activiteiten van onze neuronen uiteindelijk
weer veroorzaakt door de Big Bang en is ons bewustzijn een
epifenomeen van het bestaan van het heelal. Dat is geen vruchtbare
aanpak.
Een specifieke moderne uitwerking van het vorige misverstand
gaat er van uit dat bewustzijn geen evolutionaire functie kan
hebben omdat we ons wezens voor zouden kunnen stellen (zombies) die
kunnen overleven zonder bewustzijn. Searle stelt hier tegenover dat
men zich wellicht vogels kan voorstellen die vliegen zonder
vleugels, maar dat dit gedachtenexperiment van dezelfde
onverifieerbare waardeloosheid is. Gedachtenexperimenten zijn geen
bewijzen en zijn zelfs vaak zeer misleidend en daarmee waardeloos
omdat we ons nu eenmaal veel meer kunnen voorstellen dan in de
wereld mogelijk is. Onzichtbare mensen, bijvoorbeeld. Bewustzijn is
niet separeerbaar van ons bewuste gedrag en het bewust lopen,
zitten, eten, drinken, paren, opvoeden en dergelijke, heeft
overlevingswaarde, dus heeft bewustzijn overlevingswaarde. Als er
ooit zombies hebben bestaan dan hebben ze niet overleefd en wie
door een hersenbloeding zijn (WC-) bewustzijn verliest is niet
zelfstandig levensvatbaar.
Een andere vraag is natuurlijk of je ongestraft bepaalde bewuste
vermogens kan verliezen. Wie blind is heeft geen bewuste visuele
waarnemingen, maar kan wel bewust denken. Het is echter gevaarlijk
van daar uit te extrapoleren naar de situatie van iemand die
helemaal geen bewuste waarnemingen van welke aard dan ook zou
kunnen hebben. Dat geval kennen we in de praktijk alleen als coma.
Wat je je via gedachtenexperimenten of films kunt voorstellen zegt
niets over het leven. Een mens van mierenmaat bijvoorbeeld kan niet
bestaan (Went, 1968). De voorstelbaarheid van wandelende skeletten
of denkende spoken toont niet aan dat de spieren en huid niet nodig
zijn voor lopen of dat de hersenen overbodig zijn voor het
lastigvallen van slapende schoonheden. Zombies, wandelende
skeletten en spoken bestaan niet en dat je ze kunt bedenken bewijst
niets.
Als we ervan uitgaan dat bewustzijn een eigenschap van het
zenuwstelsel is volgt onder andere dat we zonder evidentie voor het
tegendeel aan moeten nemen dat bomen of machines geen bewuste
ervaringen beleven. Als er expliciete evidentie zou zijn dat het
hebben van een zenuwstelsel geen noodzakelijke voorwaarde voor
bewustzijn is zouden we deze a priori aanname moeten reviseren. Die
evidentie is er echter niet. Mens en dier zijn geëvolueerd,
aanvankelijk via moleculaire evolutie, daarna via cellulaire
evolutie en vervolgens via organismale evolutie, waarbij op ieder
hoger aggregatieniveau (moleculen < cellen < celverbanden
< organismen) de evolutie op lagere niveaus een rol blijft
spelen. Tijdens deze evolutie zijn lichaam, hersenen en gedrag
evenals hun mentale uitingen de `geest' aan dezelfde type
selectiekrachten onderhevig geweest: Wat fitness verhoogt kan
blijven (dat hoeft niet altijd), wat fitness verlaagt verdwijnt
(soms snel, soms op langere termijn) met de bezitters van die
kenmerken. Zenuwstelsels van alle dieren (die er een hebben) zijn
opgebouwd uit dezelfde componenten: neuronen (zenuwcellen) in
netwerken geschakeld via cel-op-cel overgangsplaatsen, de synapsen.
De membraanprocessen van neuronen en neurotransmitters (cel-op-cel
boodschapperstoffen) zijn vroeg in de evolutie ontstaan en
universeel voor alle dieren met zenuwstelsels. Kortom qua
componenten en schakelingen zijn er geen opvallende verschillen
tussen, zeg, aardwormen en mensen. De verschillen zijn voornamelijk
kwantitatief van aard: van enkele duizenden neuronen bij sommige
diersoorten tot 1012 neuronen bij de grotere
zoogdieren, zoals walvissen. Dit leidt tot de redelijke hypothese
dat bewustzijn algemener moet voorkomen in het dierenrijk dan
alleen bij één primatensoort.
Het lijkt vanuit evolutionair standpunt niet onwaarschijnlijk
dat de mate van bewustzijn gradueel verandert met het aantal
zenuwcellen, zoals dat ook geldt voor visuele waarneming, reuk,
grijpen en dergelijke. Alleen als er een alles-of-niets wet voor
bewustzijn zou gelden (bijvoorbeeld: meer dan
1010 zenuwcellen geeft bewustzijn, minder niet),
moet het a priori idee van dierlijk bewustzijn worden aangepast.
Ook daarvoor is geen evidentie. Vanuit biologisch oogpunt is daarom
de nulhypothese dat bewustzijn overlevingswaarde heeft, een
eigenschap is van zenuwstelsels en dus waarschijnlijk in diverse
afstammingslijnen in variable mate aanwezig. De meeste mensen
buiten de wetenschap gaan er intuïtief van uit dat de mate van
complexiteit van het zenuwstelsel correleert met de mate van
bewustzijn. Enquêtes (Rasmussen et al, 1993) tonen dit
duidelijk aan. Dat bewijst niet dat de nulhypothese juist is, maar
wel dat een alternatief zoals het idee dat alleen de mens
bewustzijn kent, expliciete evidentie nodig heeft om serieus te
kunnen worden genomen. Losse pols redenaties voldoen hier niet.
Als we zeggen dat bewustzijn veroorzaakt wordt door
neuronale activiteit nemen we een dualistisch standpunt in en gaan
we er kennelijk van uit dat bewustzijn iets anders zou zijn dan
hersenprocessen, terwijl de juiste visie is dat ze identiek moeten
zijn (identiteitstheorie). Deze stelling ziet Searle ook als een
akelig misverstand dat de voortgang van het onderzoek onnodig in de
weg staat. Vastheid en vloeibaarheid worden veroorzaakt door het
gedrag van de moleculen van de stof, maar vormen desondanks geen
aparte `stof' waarvan we ons af gaan vragen hoe die zich verhoudt
tot of hoe die interageert met de moleculen. Er is ook in de
natuurkunde een vorm van eigenschapsdualisme nodig. De vastheid en
vloeibaarheid zijn emergente eigenschappen van de moleculaire
interacties, worden erdoor veroorzaakt, maar hebben eigenschappen
die de microstructuur nu eenmaal niet heeft. Een molecuul heeft
geen hardheid of vloeibaarheid. Zo ook impliceert het feit dat
bewustzijn door de werking van vele neuronen wordt veroorzaakt geen
dualisme in Cartesiaanse zin. De eigenschappen van de
macroverschijnselen zijn van andere aard en daar is niets mystieks
aan.
Ook het omgekeerde misverstand komt volgens Searle voor:
Wetenschap is reductionistisch, dus als we bewustzijn verklaard
hebben in termen van neuronale activiteit is er alleen nog dat over
en hebben we het begrip bewustzijn geëlimineerd. Dit is op
dezelfde manier onjuist als wanneer men zou stellen dat het begrip
warmte geëlimineerd is nu we weten dat dit moleculaire
bewegingen beschrijft, dat hardheid en vloeibaarheid obsolete
begrippen zijn omdat we microscopische verklaringen hebben. Niets
is minder waar. Op ons normale macroscopische leefniveau blijven de
macroscopische verschijnselen van hetzelfde belang en werken we er
op dezelfde manier mee. Als een metalen bal net in een metalen ring
blijft liggen en je verhit de ring valt de bal er door. De
microscopische verschijnselen zijn daarbij niet van direct belang,
want het zijn juist de macroscopische vormen van ring en bal en hun
maten die dit verschijnsel adequaat verklaren. Bewuste ervaringen
blijven wat ze zijn en houden hetzelfde belang voor ons gedrag als
we weten dat ze het gevolg zijn van die en die activiteit in dat
hersengebied. Er worden geen begrippen geëlimineerd maar juist
toegelicht, beter hanteerbaar gemaakt, met modellen op kleinere
afmetingschalen.
Tenslotte bespreekt Searle het misverstand dat een
wetenschappelijke theorie over bewustzijn een theorie over
informatieverwerking moet zijn. Hij heeft in veel van zijn eerder
werk al betoogd dat vrijwel alles informatieverwerking kan zijn in
de ogen van een externe waarnemer die symboolwaarde toekent
aan systeemtoestanden en de verandering van toestanden dan
symboolmanipulatie of informatieverwerking noemt. De computer
verwerkt informatie omdat wij aan zijn interne toestanden
symboolwaarde toekennen. Merkwaardig veel denkers en wetenschappers
hebben de stelling verdedigd, dat andere dieren dan de mens geen
bewustzijn hebben, maar computers wel (nu of te zijner tijd). Ze
baseren zich daarbij op de analogie van symboolprocessen bij
computers en mens en zien in veel gevallen menselijke taal (of
vergelijkbare symboolprocessen) als voorwaarde voor bewustzijn.
Een probleem is dat de symboolprocessen in machines er door de
mens naar het model van een eigen vaardigheid (taal en wiskunde) in
zijn gestopt en dat er geen enkele aanwijzing is voor het idee dat
bewustzijn exclusief met taal en verwante symboolsystemen te maken
heeft. In tegendeel, als we ons bewust zijn van een stemming
(blijdschap, een rot humeur) heeft dat zelden met enig taalproces
te maken. Kleurervaringen en schoonheidservaringen hebben zelden of
nooit met taal te maken. Waarom zouden niet-sprekende dieren die
ervaringen niet hebben en niet zo nu en dan bewust het gevoel van
verliefdheid ervaren? Dissociatie van taal en bewustzijn blijkt
mogelijk, bijvoorbeeld tijdens bepaalde epileptische aanvallen in
de taalcortex. Sommige patiënten die hierdoor zowel hun spraak
als taaldenken en schrijfvermogen tijdelijk kwijt zijn kunnen
gedurende die aanvallen bewust doelgerichte handelingen uitvoeren,
zoals de weg zoeken en vinden naar een hotel in een vreemde stad
(Lecours & Joanette, 1980; zie ook Milner & Rugg, 1992).
Kortom verklaringen van bewustzijn moeten niet exclusief aan
taalprocessen worden opgehangen en zolang er geen verliefde
computers zijn evenmin aan de computermetafoor. De beste aanpak
lijkt een evolutionair perspectief bij de studie van de samenhang
tussen zenuwstelsel en gedrag, dus cognitieve en emotieve
neurowetenschap.
In de moderne neurowetenschap wordt relatief weinig over
theoretische achtergronden of filosofische aspecten van begrippen
als bewustzijn, attentie, vrije wil en dergelijke geschreven. Men
gaat ervan uit dat er een onderscheid gemaakt moet worden tussen
bewuste en niet-bewuste processen voor zowel waarnemen, handelen,
als herinneren en denken. Het ligt zodoende voor de hand te zoeken
naar verschillen tussen hersenprocessen die wel en die niet met
bewuste ervaringen correleren. Men spreekt dan wel van het
`neuronaal correlaat van bewustzijn', waarmee wordt aangegeven dat
dit niet het hele brein is, maar slechts een bescheiden deel. De
vraag is vervolgens of men deze speciale neuronale schakelingen kan
localiseren.
Het neuronaal correlaat van bewustzijn zou in principe diffuus
verdeeld kunnen zijn over het hele brein. In dat geval zou men
kunnen verwachten dat activiteiten van bepaalde herkenbare celtypen
of celnetwerken die overal te vinden zijn correleren met
bewustzijn. Voor het hersenonderzoek maakt het niet echt veel uit
of het neuronaal correlaat van bewustzijn zetelt in zulke
verspreide maar herkenbare cellen, of juist gelokaliseerd is in
één of meer netwerkjes die op één plek
bijeen liggen. Het lijkt er tegenwoordig echter sterk op dat het
neuronaal correlaat van bewustzijn goed lokaliseerbaar is in een
beperkt aantal hersengebieden, mogelijk zelfs in maar
één. Dit is interessant omdat na definitieve
lokalisatie goed kan worden nagegaan of de cellen in het
betreffende deel van het brein een andere vorm, een andere
chemische samenstelling of een ander activiteitspatroon vertonen
dan de cellen elders.
Als inderdaad slechts een bescheiden deel van de activiteit in
ons zenuwstelsel correleert met bewuste ervaring heeft dit ook
grote implicaties voor onze opvattingen over aan- of afwezigheid
van bewustzijn bij andere dieren. Alleen wanneer de betreffende
cellen en netwerken fysisch, schakeltechnisch of chemisch uniek
blijken voor de mens, moet men aannemen dat het menselijk brein het
enige is dat bewuste ervaringen ondersteunt. Localisatie op zich
zegt niet veel, omdat in de loop van de evolutie in verschillende
afstammingslijnen allerlei schakelingen naar andere delen van het
brein zijn gemigreerd. Men mag dus zeker niet stellen dat als het
neuronaal correlaat van bewustzijn bij ons in de prefrontaal cortex
zetelt dat dieren die geen prefrontaal cortex hebben dus
geen bewustzijn hebben!
Provocerende ideeën over de neurowetenschappelijke aanpak
van bewustzijnsproblemen werden door Crick en Koch naar voren
gebracht (1990, 1995) en velen hebben daaruit moed geput. Ze zijn
gewoon begonnen aan dit soort onderzoek, zonder zich langer te
laten remmen door de overweldigende en intimiderende literatuur
over bewustzijn. Hoewel Crick en Koch (1998) inmiddels aanzienlijk
genuanceerder over het onderwerp schrijven dan een decennium
geleden, is het feitelijke onderzoek mede door hun stormramacties
goed op gang gekomen. Hun eerste uitgangspunt was het volgende. Als
er processen gevonden kunnen worden die zowel niet-bewust als
bewust kunnen verlopen kan men bewustzijn als experimentele
variabele gebruiken en zien wat het verschil in
neurowetenschappelijke termen is tussen vergelijkbare processen
mèt en zónder bewuste ervaring.
Een voorlopig en toetsbaar tweede uitgangspunt kan zijn dat alle
vormen van bewustzijn één soort of een klein aantal
soorten neuronale mechanismen betreffen en dat we het verschil
daartussen en tussen niet-bewuste neuronale processen uiteindelijk
zullen kunnen meten. Als we één vorm van bewustzijn
begrijpen, begrijpen we er zodoende meer of begrijpen we ze
mogelijk zelfs allemaal. Omdat er van het zien zoveel bekend is en
omdat de mens een echt kijkdier is kunnen we het best beginnen met
het verschil tussen bewust en niet-bewust zien. Taal kan bewustzijn
wel verrijken maar is er niet essentieel voor. We gaan voorlopig
niet in op de vraag welke diersoorten wel of geen bewustzijn hebben
al nemen we aan dat er een gradueel verband tussen bewustzijn en
complexiteit van het zenuwstelsel is. We gaan er evenmin op in of
delen van het zenuwstelsel al of niet een eigen bewustzijn kunnen
hebben, stelden Crick en Koch.
De biologische rol van visueel bewustzijn is het expliciet maken
van de beste interpretatie van visuele scènes in het licht
van vroegere ervaringen van onszelf en van voorouders (via
genetische overerving van neuronale schakelingen). Hierbij hoort
een visueel werkgeheugen dat de gegevens gedurende voldoende lange
tijd beschikbaar houdt voor hersendelen die wikken en wegen en
gewilde motoracties plannen. Het nut van ongedeeld bewustzijn is
het voorkómen van aarzelingen en voorkómen van het
tegelijkertijd in gang zetten van meerdere incompatibele acties
(bijvoorbeeld tegelijkertijd gaan staan en zitten). Daarom lijkt de
verwachting van één locatie voor het neuronale
correlaat van bewust zien redelijk. Het ligt dan voor de hand te
vermoeden dat dit in de buurt van de neuronale module ligt die het
visuele werkgeheugen belichaamt. Het visuele werkgeheugen ligt
volgens werk van Goldman-Rakic (1992) in de prefrontale cortex. Dit
past bij de hypothese van Crick en Koch dat het neuronaal correlaat
van bewuste visuele perceptie in de prefrontale cortex zetelt.
Dit zijn voor een flink deel moedige of misschien overmoedige
hypothesen, maar er is inmiddels wel de nodige evidentie voor zoals
we in 8.5 zullen zien. Deze pragmatische aanpak is zeer vruchtbaar
gebleken. Aanvankelijk is er nogal wat kritiek op de aanpak van
Crick en Koch geformuleerd, wat verhelderend heeft gewerkt, maar
vervolgens is er vooral ook goed onderzoek op gang gekomen met het
doel de hypothesen te falsifiëren of ondersteunen. Dat heeft
een rijke oogst aan interessante nieuwe bevindingen opgeleverd (zie
8.5). Laten we echter de historische weg volgen en eerst kijken hoe
de oudere pogingen verliepen om hersenen en bewustzijn met elkaar
in verband te brengen (9.4) en dan ingaan op het bewuste zien
(9.5).
Al in de vijftiger en zestiger jaren waren er neurofysiologen en
neurochirurgen die probeerden een brug te slaan tussen mentale
processen zoals bewuste waarneming en herinnering enerzijds en
aanwijsbare neuronale processen anderzijds. Dit gebeurde dan
tijdens hersenoperaties aan wakkere patiënten die aan de
ingelaste proeven vrijwillig hun medewerking verleenden. De eerste
bevindingen waren meteen vrij sensationeel. Elektrische prikkeling
van bepaalde hersengebieden bleek soms filmachtige herinneringen op
te roepen, compleet met de ervaring van de patiënt dat het
echt gebeurd was of op dat moment echt gebeurde. Helaas was er op
de technieken nogal wat aan te merken en de prikkels waren
waarschijnlijk erg sterk en slecht gedoseerd, zodat enorme delen
van het brein op onnatuurlijke wijze werden geactiveerd. Naarmate
de technieken beter werden en de prikkeling localer en beter
gedoseerd kon worden toegediend bleken de rapporten van
patiënten minder bloemrijk en uitvoerig. Er bleven meestal
slechts vrij bescheiden ervaringen over, zoals het zien van een
enkel object, het horen van een woord of kreet, de herinnering van
een geur. Het meeste opzien baarde vervolgens het werk van Benjamin
Libet en sterk vereenvoudigde versies van zijn vergaande
interpretaties van de bevindingen hebben hun weg gevonden naar de
populaire en de filosofische literatuur. Omdat er nogal wat vreemde
en ongefundeerde ideeën op zijn gebaseerd is het de moeite
waard op dit werk nader in te gaan en kort aan te geven waarom de
oorspronkelijke interpretaties niet al te serieus meer moeten
worden genomen.
Libet concludeerde uit experimenten, die we zo zullen
beschrijven, het volgende: (1) Het duurt vrij lang, tot wel een
halve seconde, voor een zintuiglijke prikkel tot het bewustzijn
doordringt; (2) Het bewustzijn refereert de ervaring dan diezelfde
wachttijd terug in de tijd, zodat de bewuste ervaring lijkt te
komen op het moment van prikkeling; (3) Een ge`wilde' beweging
wordt al zo'n halve seconde voor de start van de uitvoering in het
zenuwstelsel opgewekt en pas zo'n 350 ms later ervaren we bewust de
`wil' om de beweging uit te voeren; (4) Hoewel de vrije wil dus
(volgens Libet) schijn is en de beweging in feite onbewust wordt
opgewekt, is er nog op het laatste nippertje een door de wil
gegeven veto mogelijk, dat kan voorkomen dat de beweging wordt
uitgevoerd.
Het is logisch dat dit soort stellingen voer voor bespiegelingen
vormen. Sommigen concludeerden uit Libet's werk dat geestelijke
verschijnselen zich in een niet-fysisch domein afspelen en daardoor
vrijelijk in tijd konden worden verschoven, of tenminste dat Libet
had aangetoond dat geest en hersenen onafhankelijke processen zijn.
Ook de vrije wil werd met grote overgave naar de prullenbak
verwezen. Alle acties gebeuren volgens Libet immers spontaan vanuit
de niet-bewuste delen van het zenuwstelsel en worden nog maar op
het nippertje van een bewust labeltje voorzien dat aangeeft dat de
eigenaar van de hersenen het er mee eens is. Het werk van Libet en
zijn groep is natuurlijk uitgebreid bekritiseerd, onder andere door
aanhangers van de identiteitstheorie, maar een echt grondige
kritiek vanuit de meer neurofysiologische hoek is helaas lang
uitgebleven. Het beste voldoet wellicht het recente artikel van
Gomes (1998), waarin zeer systematisch wordt aangetoond dat alle
vier de boven gegeven conclusies uit het werk van Libet
ongefundeerd zijn. Het is daarmee niet bewezen dat ze onjuist zijn,
maar ze volgen in elk geval geen van alle uit het experimentele
werk van Libet en medewerkers, noch uit dat van andere
hersenonderzoekers. Het lijkt ons belangrijk genoeg om hier wat
preciezer op in te gaan, maar voor de details verwijzen we naar
Gomes (1998).
Allereerst moet worden benadrukt, dat de kritiek zich niet richt
op de resultaten van Libet en zijn groep, maar op hun
interpretaties. De meeste meetresultaten zijn vergaard in de
periode 1965-1985 en de beste overzichten zijn Libet (1985, 1993).
De eerste vraag was: Hoe meten we het moment waarop een waarneming
bewust wordt? Libet prikkelde de somatosensorische cortex met een
reeks korte pulsjes waarvan hij de prikkelfrequentie f (aantal
pulsjes per seconde), de amplitude van de stroomsterkte per puls A
(in mA) en de totale duur T (in ms) kon instellen. Fig. 1A
geeft een voorbeeld van zo'n `pulstreintje' van T sec duur. Om zo
een bewuste waarneming op te roepen, bijvoorbeeld het gevoel van
aanraking van de linkerarm, moest de frequentie in elk geval boven
de 15 pulsjes/seconde zijn. Libet bepaalde de waarde van
pulstreinduur T nodig om bij minimale amplitude
A=Amin nog net een bewuste waarneming te krijgen
en vond hiervoor een T-waarde van minimaal 500 ms (0,5
seconde).
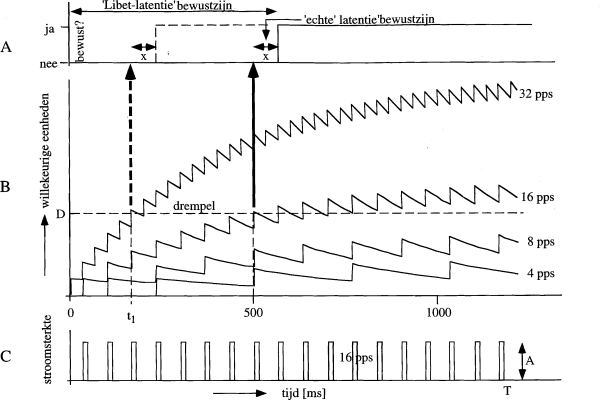
Figuur 1: In A is gesymboliseerd hoe het neuronaal correlaat van
bewustzijn al (`ja') of niet (`nee') actief kan zijn. Het wordt
aangestuurd door een netwerk waarvan de activiteit in B is
geïllustreerd. Dat netwerk omvat onderdrempelige sommatie en
het stuurt pas een prikkel naar het neuronale correlaat van
bewustzijn zodra drempel D is overschreden. Dit moment van
drempeloverschrijding is met een vetgedrukte verticale pijl
aangegeven. Vanaf dat moment duurt het nog x ms voor het neuronaal
correlaat van bewustzijn reageert. We zien de onderdrempelige
sommatiepotentialen voor vier gevallen, namelijk prikkeling met 4,
8, 16 en 32 pps (=pulsen per seconde). In C is de electrische
prikkeling met pulstreintjes gesymboliseerd, waarbij hier het geval
16 pps als voorbeeld is genomen. De amplitude van de prikkel is A,
de duur T. Verdere toelichting wordt in de tekst gegeven.
Nu komt het interpretatieprobleem. Libet ging ervan uit dat deze
pulstreinduur aangeeft hoe lang het duurt voor de trein tot
bewustzijn leidt. Hij zag deze duur dus als een soort latentie van
het bewustzijn en leidde er uit af dat het minstens een halve
seconde duurt voor een prikkeling tot het bewustzijn doordringt.
Dat is natuurlijk nogal slordig geredeneerd! Bij neuronale
prikkeling moet men normaal gesproken door sommatie van
onderdrempelige signalen een drempel overschrijden,
vóór er gereageerd wordt. Dan geldt voor deze uiterst
zwakke prikkeling (minimale amplitude) waarschijnlijk gewoon dat
het grootste deel van de pulstrein onderdrempelig is. In figuur 1B
is aangegeven hoe de onderdrempelige sommatie zou kunnen
plaatsvinden en hoe dan pas na (bijvoorbeeld) 500 ms de drempel
wordt overschreden van een netwerk dat vooraf gaat aan het
neuronale correlaat van bewustzijn. Fig. 1B geeft dus steeds
de activiteit in dat niet-bewuste netwerk, terwijl de dikke pijlen
aangeven wanneer de drempel van dat netwerk wordt overschreden en
er dus een signaal naar het neuronaal correlaat van het bewustzijn
gaat.
Bekijken we het signaal dat hoort bij een stimulus van 16 pps in
Fig. 1B dan geeft de verticale pijl aan wanneer het `neuronaal
correlaat van bewustzijn' wordt geprikkeld en daarna duurt het
(bijvoorbeeld) nog x ms voor die reageert. Zodoende duurt het 500+x
ms vanaf de start van de pulstrein tot de bewuste ervaring en dat
hebben we in Fig. 1A de `Libet-latentie' van bewustzijn
genoemd. De `echte' latentie van het neuronaal correlaat van het
bewustzijn is vanaf het moment dat het een prikkel van het
voorafgaande netwerk ontvangt slechts x ms. Dat zou dan betekenen,
dat het bewustzijn helemaal geen grote latentie heeft (hier x ms),
mits het voorafgaande corticale netwerk bovendrempelig wordt
aangestuurd. De `echte' reactietijd x kunnen we niet uit de
metingen van Libet afleiden, zodat zijn metingen niets zeggen over
de latentie van bewuste ervaringen.
Ons (Gomes') alternatieve model van onderdrempelige sommatie
klopt overigens uitstekend met Libet's meetresultaten, want voor
sterkere prikkeling (grotere amplituden) vond Libet dat de
pulstrein aanzienlijk korter kan zijn om tot bewuste ervaringen te
leiden. Sterkere prikkels leiden tot sneller bereiken van de
drempel. Het sommatie-tot-drempel-model maakt ook duidelijk waarom
de frequentie van de pulstrein niet te laag mag zijn. Dan immers
lekt tussen twee pulsen tenminste net zo veel activatie weg als er
per puls wordt opgewekt. Dit is in Fig. 1B geïllustreerd
voor de signalen horend bij de lagere prikkelfrekwenties 8 en 4
pps. Bij hogere prikkelfrekwenties, zoals de 32 pps uit
Fig. 1B, zien we dat de drempel al eerder wordt overschreden,
namelijk t1 ms na de start van de stimulus. Kortom, stelling 1 van
Libet (het duurt minstens 0,5 sec voor een prikkel bewust wordt) is
niet verdedigbaar op grond van zijn meetresultaten.
Laten we vervolgens stelling 2 van Libet nader bekijken, het
terugrefereren in de tijd van de bewuste ervaring. Libet ontdekte
allereerst dat bij zo zwak mogelijke elektrische prikkeling van de
huid al na 33 tot 100 ms een bewuste ervaring optreedt. Het is in
termen van het alternatieve model (Gomes, 1998) net alsof hier
nauwelijks onderdrempelige sommatie nodig is en de prikkel na heel
korte latentie bewust wordt. Dat is ook niet zo gek, want via de
huid loopt de fysiologisch normale, dus effectieve, weg om de
cortex aan te sturen. Daarvoor kan de gevoeligheid (de verticale
stapjes in de signalen in fig.1B) veel groter zijn, zodat de
drempel al zeer snel wordt overschreden en het daarna slechts de
latentie x ms duurt voor er een bewust percept optreedt.
Nu deed Libet het volgende experiment. Hij prikkelde de huid van
bijvoorbeeld de linkerarm elektrisch en tevens in de cortex het
gebied dat correspondeert met de rechterarm. De vraag aan de
proefpersonen was in een bepaald experiment om de duur van de twee
soorten stimuli te vergelijken. Die duren bleken in de ervaring
gelijk lang te zijn als de duur van de corticale prikkel--vanaf het
moment waarop deze volgens Libet bewust wordt--gelijk is aan de
duur van de huidprikkel. Dat past dus prima in bovenstaand
alternatief model (volgens Gomes, 1998). Stel dat bij een bepaalde
prikkelsterkte de drempel corticaal na bijvoorbeeld 200 ms is
overschreden (een sterkere corticale prikkel dan de minimale die we
boven noemden). Als de prikkel daarna nog 300 ms wordt aangehouden,
dan vinden de proefpersonen hem even lang duren als een huidprikkel
van 300 ms. Het is alsof de duur van het onderdrempelig
sommatieproces niet meetelt in de bewuste ervaring. Dat zouden we
volgens ons alternatieve model precies verwachten indien het
onderdrempelig sommeren een actie is in een voorafgaand niet-bewust
netwerk. De echte bewustwordingslatentie x zou bij beide soorten
prikkels gelijkelijk het moment van optreden verschuiven en dus
onmerkbaar zijn in deze tijdsduurvergelijking.
Vervolgens deed Libet het cruciale experiment dat de meeste
verwarring heeft opgeleverd. Hij gaf de P-stimulatie, dat is de
perifere stimulatie van de huid, op een variabel tijdstip na de
start van de corticale oftewel C stimulatie en vroeg de
proefpersonen of de ene (P-gerelateerde) dan wel de andere (C
gerelateerde) waarneming eerder begon. Het bleek dat de
P-stimulatie eerder werd ervaren, zelfs als deze 450 ms na de start
van de zwakste C-stimulatie werd gegeven. Libet had verwacht dat ze
gelijk zouden lijken op te treden als ze gelijk zouden beginnen en
was dus verrast. Omdat hij ervan uitging dat beide een latentie van
bijvoorbeeld 0,5 seconde hadden, moest dit betekenen dat het
tijdstip van optreden van de P-stimulatie terug werd geschoven in
de waargenomen tijd! De conclusie werd dus: prikkeling van de huid
wordt ons pas na 0,5 sec bewust maar die ervaring wordt in onze
perceptuele tijd teruggeschoven naar het moment van prikkeling.
Voer voor mystici. In feite is dit gewoon onzuiver redeneren!
Ons alternatief (neurofysiologisch plausibel) model zegt
simpelweg dat de C-stimulatie pas tegen het eind van de pulstrein
de sommatiedrempel bereikt en dan pas de bewuste corticale module
aanstuurt. Een P-stimulus die op dat moment van
drempeloverschrijding gegeven wordt zal er dus gelijktijdig mee
lijken. Zowel P als C stimulus zouden dan de echte latentie tot
bewustworden van x ms hebben, maar deze echte latentie gaat voor de
C-stimulus pas in als de drempel overschreden wordt, als er
voldoende opeenvolgende pulseffecten zijn gesommeerd, in een
voorafgaande sommatieschakeling. Weg mysterie van tijdgoochelen! We
verwijzen naar Gomes (1998) voor een veel uitgebreidere
behandeling, waarbij ook de zogenaamde `maskerings' experimenten
van Libet worden besproken.
De derde stelling van Libet, dat vrije wil een illusie is en dat
een `wilsbesluit' dus niet aan het begin van een causale keten
staat die tot een actie van het organisme leidt, is wellicht het
meest bejubeld, vooral door behavioristisch ingestelde filosofen en
psychologen. Is dat echter wel zo geloofwaardig aangetoond? Ons
speurwerk begint nu bij de `Bereitschaftspotential' die door
Kornhuber en Deecke (1964) werd ontdekt en door Libet onder de naam
`Readiness Potential' werd omarmd. Een proefpersoon wordt gevraagd
om steeds weer een wijsvinger te bewegen zodra hij of zij daar `zin
in heeft', het dus bewust `wil'. Het EEG (electro encephalogram)
van de proefpersoon wordt continu gemeten. Na een flink aantal
vingerbewegingen gingen de onderzoekers terugzoeken (`reverse
correlation') hoe de EEG signalen eruit zagen direct voorafgaand
aan de vingerbeweging. Wat bleek als je er minstens zo'n 20
middelt? (Als je dat niet doet zie je niets dan ruis). Ongeveer 90
ms voor de beweging is er een `premotorpotentiaal' te zien
(Fig. 2), maar al ongeveer 750-800 ms voor de beweging begint
treedt er een EEG-golf op die Kornhuber en Deecke dus
`Bereitschaftspotential' noemden: Fig. 2.
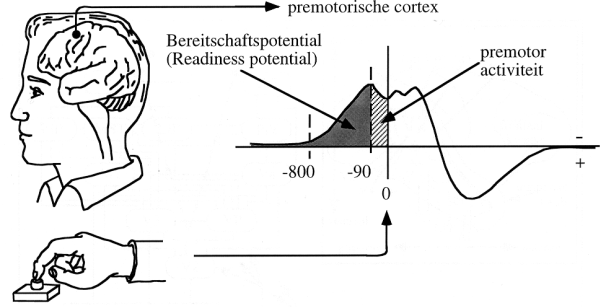
Figuur 2: (Vrij naar Kandel en Schwartz, 1992). Als een
proefpersoon op willekeurige momenten een knop indrukt en men
middelt electronisch het electro-encefalogram (EEG) voorafgaande
aan (en volgend op) een stuk of 20 vingerbewegingen, dan vindt men
een golfvorm als hier rechts geschetst. Al 90 ms voor de
vingerbeweging is de premotor cortex actief, die de motorcortex
aanstuurt, van waaruit de spieren van de vinger worden bediend.
Voorafgaand aan die premotor activiteit werd echter een golfvorm
gevonden die al zo'n 800 ms voorafgaand aan de actie begint. Dit
noemt men de `Bereitschaftspotential'. In de tekst wordt uitgelegd,
waarom deze `Readiness potential' niet bewijst dat de beslissing om
een vingerbeweging te maken al ruim voor men er zich van bewust
wordt is genomen. De Bereitschaftspotential bewijst dus niet dat
handelingen buiten de `wil' om gaan of dat er geen vrije wil zou
bestaan.
Het bewuste `willen' van de vingerbeweging leek echter korter
voor de beweging op te treden dan 750-800 ms. (We laten een
beschrijving van de experimenten die dit moesten aantonen
achterwege). Daarom vonden velen dat de conclusie voor de hand
ligt, dat de beslissing onbewust wordt genomen (al 800 ms voor de
vingerbeweging) en dat men zich pas later van die beslissing bewust
wordt, waarbij het `aanvoelt' alsof het een eigen wilsbesluit zou
zijn geweest. Vrije wil is daarmee een illusie, beslissingen om te
bewegen worden onbewust genomen en dan lijkt het alleen maar alsof
we ze wilden. Het probleem met deze conclusie wordt duidelijk als
men probeert zich in de situatie van de betreffende proefpersonen
te verplaatsen (Gomes, 1998).
Introspectief lijkt het er helemaal niet op alsof men voor
iedere vingerbeweging een apart wilsbesluit neemt. Integendeel, men
neemt het bewuste besluit aan het begin van het hele experiment om
op random ogenblikken een vingerbeweging te maken. Dat stelt
mogelijk in het zenuwstelsel een random generator in werking die
starttijdstippen opwekt en dan op die tijdstippen de
vingerbeweegprogrammering start. Je hoeft je alleen van de ophanden
zijnde beweging bewust te worden op een laat tijdstip om nog een
veto te kunnen geven en de experimenten van Libet toonden aan dat
je dat ook kunt. Het is dus zeer wel denkbaar dat het wilsbesluit
inderdaad niet per vingerbeweging wordt genomen, maar van meer
strategische aard is. Je besluit bewust aan het experiment mee te
doen en zet daarmee een complex hersenprogramma in beweging dat
voor de rest zorgt, inclusief de willekeurige beweegmomenten. Dit
ge`wilde' globale programma zorgt voor de uitvoering van het hele
spel en is bij de uitvoering slaafs genoeg om voor iedere
vingerbeweging nog even te controleren of we hem echt wel willen
door hem aan te melden bij de bewustzijnsmodule. We zien hier een
houdbare alternatieve opvatting van de werking van de vrije wil,
die meer stuurt op hoofdlijnen. Daarmee verdwijnt de paradox dat we
een gewilde beweging in feite onbewust starten en dan achteraf de
illusie krijgen dat we de beweging bewust wilden.
Het probleem van de vrije wil, illusie of niet, is hiermee
natuurlijk niet opgelost. Wel is dit voorbeeld mogelijk voldoende
om te laten zien dat men erg voorzichtig moet zijn met het trekken
van harde conclusies uit dit soort experimenten. De interpretatie
hangt sterk af van het gehanteerde model. Als we vrije wil zien als
de innerlijke ervaring die gepaard gaat met het nemen van globale
beslissingen om zo-en-zo te handelen of om die-en-die strategie te
volgen, dan is het de mentale kant van de neuronale opdrachten om
bepaalde hersenprogramma's te starten, te stoppen of te maken
(programmeren). We zijn ons lang niet van alle en zelfs niet van
vele detailbeslissingen bewust. Tijdens automatisch handelen neemt
het zenuwstelsel allerlei beslissingen waarvan we ons niet bewust
zijn. Slechts de start van een heel gedragsprogramma en enkele
controlemomenten tijdens de uitvoering hoeven tot het bewustzijn
door te dringen, respectievelijk er uit voort te komen.
Met `vrije wil' kan dus bijvoorbeeld bedoeld worden dat
rapporteerbare interne afwegingen die principieel meerdere
mogelijke uitkomsten kunnen opleveren voorafgaan aan de start van
het gedragsprogramma, dat de gekozen uitkomst implementeert of een
stap naderbij brengt. Er is hierbij uiteraard geen sprake van
ultieme vrijheid om te willen, want alle interne afwegingen
berusten steeds op interne gegevens en structuren die al aanwezig
waren in het brein van degene die aan het afwegen is, in combinatie
met de ervaring door die persoon van externe gebeurtenissen. Ruis
en chaosprocessen kunnen de voorspelbaarheid van een wilsbesluit
erg klein maken, zelfs voor degene die `wil', maar
onvoorspelbaarheid is niet hetzelfde als ultieme vrijheid. Het
begrip `vrije wil' lijkt ons vrijwel uitsluitend bruikbaar in het
geval waarbij we hier en nu kijken of de beslissing op grond van
rapporteerbare afwegingen vooraf gaat aan en de oorzaak is van het
volgende gedragsprogramma.
Het bijzondere van de beslissingen die wel bewust worden ervaren
is dat ze met prioriteitstellingen te maken hebben. Als je
autorijdt en tegelijk met een passagier een gesprek voert gaat het
besturen van de auto buiten het bewustzijn om. Het zenuwstelsel
neemt autonoom (onbewust) beslissingen over schakelen, gaspedaal
bedienen, aan het stuur draaien en dergelijke. Als je ineens gevaar
bemerkt wordt er als het ware een interrupt gegeven, het gesprek
stokt, de aandacht gaat naar het autorijden en je neemt bewuste
beslissingen. Iemand die bijvoorbeeld tennist voert evenmin
voortdurend ge`wilde' bewegingen uit. De meeste bewegingen zijn
daarbij automatismen of reflexen. De vrije wil uit zich in
gedachten over de strategie. Laat ik een effectbal proberen, nu
moet ik smashen, enzovoorts. Gelukkig ben je je dan niet bewust van
de uitvoeringsdetails, zoals de buighoek van ellebooggewricht of
pols. Slechts een fractie van de activiteiten van het zenuwstelsel
dringt door tot het bewustzijn en die fractie heeft steeds met
belangrijke zaken te maken, met hoofdlijnen van de
keuzemogelijkheden en beslissingen, met prioriteitstellingen.
De `Bereitschaftspotential' heeft in deze opvatting niets met
het onderscheid tussen gewilde (bewuste) en niet-bewuste
handelingen te maken. Keller en Heckhausen (1990) lieten zien dat
ook niet-bewuste motorische acties door dezelfde soort
Bereitschaftspotential vooraf worden gegaan. De vrije wil richt
zich vermoedelijk niet gauw op zulke details als het maken van een
buigbeweging met een vinger! De vrije wil zal eerder te maken
hebben met de keuze mee te doen aan het experiment en de opdracht
aan het zenuwstelsel op random momenten een buigbeweging te maken.
Verder houdt het bewuste deel van het zenuwstelsel dan in de gaten
of de buigbewegingen ook plaats vinden en of ze op het automatisch
geprogrammeerde moment nog wel wenselijk zijn.
De alternatieven die we besproken hebben laten zien dat de
conclusies van Libet niet volgen uit de experimentele resultaten
(al kunnen ze altijd nog toevallig waar zijn). Echter onze
alternatieven hebben ook bedenkelijke kanten. We hebben net gedaan
of er een aparte corticale module is waarin het bewustzijn van de
waarneming huist en een `vrije-wil' module die beslissingen neemt
over strategieën en globale handelingspatronen. Dat lijkt
verdacht veel op het soort model dat Descartes bedacht, waarbij een
geest waarneemt wat er in het sensorium (de Cartesische schouwburg)
gebeurt en het motorium bespeelt als een soort poppenspeler zijn
poppentheater. De niet-materiële geest van Descartes is nu
weliswaar vervangen door concrete hersenmodules voor waarneming en
voor actiesturing, maar is dat nu zo'n geweldige conceptuele stap
vooruit? Wij vermoeden van wel, omdat er inderdaad evidentie is
voor de aanname van specifieke modules die met bewuste ervaringen
te maken hebben (zie 8.6), terwijl de meeste andere modules van het
brein hun taken verrichten zonder dat we ons daarvan bewust
worden.
Als bewustzijn lokaliseerbaar is, is de eerste stap gezet naar
het bestuderen van de mechanismen die eraan ten grondslag liggen.
Trouwens, welke alternatieven zijn er bedacht? Als je het
Cartesiaans theater afwijst (Dennett en Kinsbourne, 1992) verval je
automatisch in een soort holistisch model: bewustzijn als
gedistribueerde eigenschap van het gehele brein. Daar spreekt erg
veel tegen (Baars, 1998), zoals we nog zullen zien. Men kan het
geheel van modules die samenhangen met bewuste ervaringen ook het
`Besturingssysteem' van het brein noemen (van de Grind, 1997), een
besturingssysteem dat de werking van een enorm groot en complex
parallel systeem met vele semi-autonome modules coördineert.
Anderen hebben een voorkeur voor metaforen als Executive System,
directiekamer, en dergelijke. De moderne opvatting van een directie
als groep medewerkers met coördinerende taken in plaats van
als goddelijk geïnspireerde leiders is in deze metafoor
analoog aan de verschuiving van Descartes `bewuste onstoffelijke
geest' naar de moderne stoffelijke bewuste hersenmodules.
Lokaliseerbaar, beschrijfbaar, tastbaar, functioneel en toch een
beetje verheven.
In Fig. 3 geven we een zeer vereenvoudigd overzichtsschema
van de banen die van cellen in het netvlies van het oog, via
tussenstations zoals de primaire visuele cortex V1, leiden naar
allerlei `hogere' hersengebieden voor visuele waarneming. De inzet
rechtsboven toont een zijaanzicht van apenhersenen (makaak), waarop
de ligging van de genoemde gebieden te zien is. Iedereen weet
natuurlijk, dat we zonder functionerende netvliezen in de ogen niet
kunnen zien, maar dan is zowel bewust als niet-bewust zien
onmogelijk, dus dit zegt niets over de localisatie van het bewuste
zien. Volgens Crick en Koch (1990, 1995) is activiteit in de
primaire visuele cortex V1 evenmin essentieel voor visueel
bewustzijn, al levert V1 normaal natuurlijk de meeste gegevens aan
het neuronaal correlaat van bewustzijn. Ze formuleerden deze
stelling omdat V1 geen directe verbindingen heeft met de frontale
cortex, die volgens lesiestudies wel essentieel lijkt voor bewust
zien. Hun hypothese is dan ook dat het neuronale correlaat voor
bewust zien in de prefrontale cortex (een associatiegebied in de
frontaalkwab) zetelt. Daar is in elk geval, zoals eerder vermeld,
ook het werkgeheugen voor posities en voor visuele objecten
gevonden, namelijk in en om de sulcus centralis, de hersengroeve
die in Fig. 3 vooraan horizontaal verloopt.
Als het voorbewerkingssysteem tot en met V1 intact is en er
beschadigingen (lesies) in één van de hogere visuele
gebiedjes optreden (V2, V3, VP, V4, enz) kan de patiënt in
algemene zin wel bewust zien, maar mist de bewuste perceptie van
die aspecten van de visuele wereld die door het ontbrekende
gebiedje worden verwerkt. Bijvoorbeeld gebiedje MT, ook wel V5
(vijfde visuele cortex) genoemd, verwerkt informatie over visuele
beweging (van niet te lage snelheden, want die worden via de
temporale cortex verwerkt, samen met vormanalyse en vorm uit
beweging). Een patiënt die dat gebiedje mist is
bewegingsblind, maar kan wel kleuren zien en benoemen, vormen zien,
objecten herkennen en dergelijke. De beroemdste patiënte met
deze lesie, een Duitse dame, is in laboratoria over de hele wereld
onderzocht met een enorm scala aan visuele tests. Ze mist alleen de
waarneming van visuele beweging, behalve dan van ultralage
snelheden. Dit is overigens een enorme handicap, want ze kan auto's
in het verkeer niet zien bewegen. Ze ziet een naderende auto klein
en ver weg en ineens duikt dezelfde auto groter en dichtbij opnieuw
op. Als ze thee inschenkt loopt het kopje altijd over, want ze ziet
het niveau in het kopje niet zoals wij geleidelijk stijgen
(bewegen). De thee in het kopje en de straal uit de kan lijken
bevroren, het theeniveau springt schoksgewijs van een laag niveau
naar een hoog niveau en voor ze kan stoppen met inschenken is alles
al over de rand. Het is natuurlijk vrij moeilijk om haar ervaringen
in woorden te beschrijven, want het is slecht mogelijk je in deze
relatief vreemde visuele handicap in te leven.
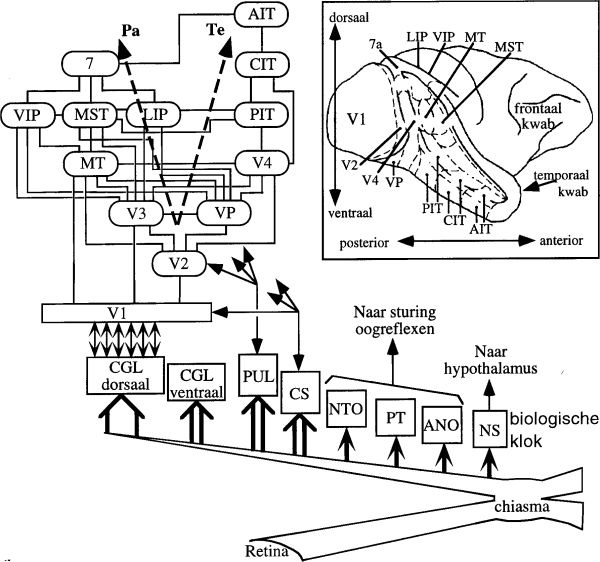
Figuur 3: Een schema dat de stations voor visuele
gegevensverwerking aangeeft die gepasseerd worden van de retina tot
het neuronaal correlaat van bewustzijn (mogelijk in de
frontaalkwab). De vele zijtakken van de tractus opticus, zoals naar
NS (nucleus suprachiasmaticus), ANO (accessorische kern van de
tractus opticus), PT (pretectum), NTO (kern van de tractus
opticus), zijn hier minder van belang. CS (colliculus superior) en
PUL (pulvinar) spelen een rol bij `blindzien' (zie tekst). De
hoofdbanen voor visuele perceptie lopen via CGL (corpus geniculatum
laterale) naar de primaire of `striate' cortex V1, waarna er aparte
`stromen zijn via de temporale cortex (baan Te) en pariëtale
cortex (baan Pa), die respectievelijk verantwoordelijk lijken voor
vormzien (`wat' is het) en ruimtelijk zien (`waar' is het). Sommige
van die hogere (`extrastriate') cortexgebiedjes komen in de tekst
ter sprake. In de inzet is een rechterzijaanzicht gegeven van de
cortex van een makaak aapje. Let ook op de horizontale groeve in de
frontaalkwab, waar het werkgeheugen gelokaliseerd lijkt. In die
buurt zoeken velen ook het neuronaal correlaat van bewustzijn,
zoals in de tekst wordt beschreven.
Bij metingen met micro-elektroden aan individuele zenuwcellen in
MT (V5) is ook gebleken, dat deze cellen zich vrijwel alle met
visuele beweging bezig houden. Een typische cel in MT reageert
optimaal op een bepaalde bewegingsrichting en snelheid van een
visueel patroon dat in het `receptieve veld' (RV) van de cel wordt
aangeboden. Iedere cel in het visuele systeem kan uitsluitend of
voornamelijk uit een zeer duidelijk begrensd gebied in de
buitenwereld (op een scherm) geprikkeld worden en dat gebied noemt
men het RV. Dat RV is zeer klein (bijvoorbeeld een halve graad
gezichtshoek) in het netvlies en wordt groter naarmate we hoger in
de hiërarchie van cortexgebieden uit Fig. 3 komen. Cellen
in MT hebben een RV van vele graden doorsnede en als ergens in hun
RV, of liefst zelfs overal, beweging optreedt van hun
voorkeursrichting en in de buurt van hun optimale snelheid dan
vuren ze veel actiepotentialen per tijdseenheid af. Bill Newsome en
medewerkers (zie bijv. Salzman et al, 1990, of Parker en Newsome,
1998) hebben bij wakkere aapjes gemeten aan de activiteit van
individuele MT cellen en groepen van deze cellen tegelijk via
andere elektroden elektrisch geprikkeld. De aapjes waren getraind
met hendeltjes of met oogbewegingen aan te geven of ze beweging
zagen en zo ja in welke richting.
Als de bewegingsrichting overeenkwam met de voorkeursrichting
van de bemeten cel vuurde de cel hard. Als de aap getraind was de
visuele attentie op het RV te richten vuurde de cel nog wat harder.
Cellen met een bepaalde voorkeursrichting en voorkeurssnelheid
hebben de neiging in clusters bijeen te liggen. De interessantste
bevindingen kwamen voort uit combinatie van gedragsproeven met
microstimulatie van zulke celclusters. Indien cellen van zo'n
cluster door de beweging van hun voorkeursrichting visueel optimaal
werden aangestuurd rapporteerde de aap inderdaad ook die
bewegingsrichting. Toen de onderzoekers vervolgens datzelfde
celcluster elektrisch prikkelden en geen visuele stimulus gaven,
rapporteerde de aap ook de betreffende bewegingsrichting te zien.
Prikkeling van die specifieke groep cellen bleek steeds voldoende
voor de rapportage door de aap dat hij de betreffende beweging zag.
Omgekeerd konden de onderzoekers de cellen ook remmen door de
stroomrichting van de elektrische prikkel om te keren en het bleek
dat de aap dan de beweging nooit rapporteerde, ook niet als deze
optimaal visueel werd aangeboden. Stilleggen van de betreffende
groep cellen was een voldoende voorwaarde om de visuele prikkel
niet te zien. Activiteit van dit cluster cellen bleek dus
noodzakelijk en voldoende te zijn voor rapporteerbare waarneming
van beweging met de bij dit cluster horende optimale
bewegingsrichting.
Dit werk toonde overtuigend aan dat er een vast (causaal)
verband is tussen bewuste perceptie (expliciete rapporteerbaarheid)
van bepaalde beweging en de activiteit van het cluster cellen in MT
dat speciaal voor die beweging gevoelig is. De experimenten zijn
door vele andere groepen inmiddels herhaald en het onderzoek richt
zich tegenwoordig meer op de invloed van attentie op de
gevoeligheid van dit soort cellen of gebieden. Globaal gesproken
blijkt attentie gericht op het gebied waar de RVn van bepaalde
cellen liggen de vuurfrequentie van die cellen te vergroten als ze
hun voorkeursprikkel aangeboden krijgen. Met het intuïtieve
idee dat cellen die het hardst roepen het best doordringen tot het
neuronaal correlaat van bewustzijn is dan een brug gelegd tussen
het niveau van individuele zenuwcellen en het expliciete
rapporteren, de bewuste visuele waarneming, van de aap.
Het zal duidelijk zijn dat er voor alle bewegingsrichtingen (zeg
om de 30 graden) aparte celclusters verantwoordelijk zijn en dat er
voor verschillende snelheidsbereiken ook verschillende celclusters
verantwoordelijk zijn. Een individuele cel heeft namelijk weliswaar
een zeer bepaalde optimale richting, maar reageert afnemend sterk
op nabijgelegen richtingen en valt terug op de
evenwichtsvuurfrequentie (ook `spontane activiteit' genoemd) als de
richting zo'n 45-90 graden van de optimale afligt. Meestal worden
de cellen geremd door richtingen in de buurt van tegengesteld aan
hun voorkeursrichting en vallen dan pas helemaal stil. Op analoge
wijze is er naast een bereik van richtingen ook een bepaald
snelheidsbereik waarvoor zo'n cel reageert. Tesamen bedekken de
cellen van MT dus het hele zichtbare gebied van snelheden (op hele
lage snelheden na, zie boven) en van bewegingsrichtingen. Er is
inmiddels zoveel onderzoek gedaan aan dit gebied MT, dat het
mogelijk is dit alles kwalitatief uitstekend en kwantitatief
redelijk in elkaar te passen.
Gebied MST (Fig. 3) is eveneens uitgebreid bestudeerd en
blijkt zich vooral met visueel-gestuurde navigatie bezig te houden.
Cellen in MST zijn gevoelig voor opzwelbeweging (divergentie) of
inkrimpbeweging (convergentie), voor rotatie rechtsom of linksom.
Dit is het soort bewegingsinformatie dat aangeeft dat we op iets af
bewegen (opzwelbeweging), er vanaf bewegen of dat we ons hoofd of
lichaam draaien om onze voortbewegingsrichting. De cellen in MST
blijken dan ook vooral een rol te spelen bij het sturen van de
ego-beweging. Ook hier is er een goede correlatie tussen waarneming
en celeigenschappen gevonden in proeven met wakkere getrainde
aapjes (Britten en van Wezel, 1998).
Alle cortexgebieden boven de primaire visuele cortex V1 hebben
te maken met bewuste ervaringen, ongeacht of je ze elektrisch
prikkelt of visueel via V1. Al deze gebiedjes hebben directe
verbindingen met de frontaalkwab en dit lijkt inderdaad (zoals
Crick en Koch postuleerden) cruciaal te zijn voor de bewustwording
van hun activiteit. V1, ook wel striate (gestreepte) cortex
genoemd, heeft geen directe banen naar de frontaalkwab. Alle andere
cortexgebiedjes in het schema noemt men wel tezamen en in contrast
tot V1, de extrastriate cortex. Het idee is dus dat de striate
cortex geen en de extrastriate cortex wel directe toegang heeft tot
het neuronaal correlaat van bewust zien. De discussie spitst zich
toe op de vraag of ieder van de extrastriate gebiedjes zelf een
neuronaal correlaat van bewust zien omvat voor wat betreft het
eigen specialisme (verdeeld bewustzijnscorrelaat) of dat ze ieder
toegang hebben tot één centraal neuronaal correlaat
van bewust zien in de frontaalkwab. Voor dit laatste model is veel
te zeggen, omdat het werkgeheugen ook in de frontaalkwab is
gelokaliseerd (Goldman-Rakic, 1992). Voor een homologe locatie van
werkgeheugen bij mensen is de laatste tijd ook veel evidentie
gevonden in hersenscanonderzoek (fMRI) bij mensen die geheugentaken
verrichten (Courtney et al, 1998). Anderzijds zijn er ook
prominente verdedigers van het multi-visuele-bewustzijn idee, zoals
Zeki en Bartels (1998).
In Fig. 3 geven de twee dikke stippellijnen aan, dat er
twee hoofdtrajecten vanuit V1 `omhoog' vertrekken, de
pariëtale (Pa-) baan die vooral met ruimtelijk gedrag te maken
heeft en de temporale (Te-) baan die vooral met vorm en
objectherkenning te maken heeft. Lesies in de temporaalkwab bij de
mens leiden dan ook tot problemen die hiermee te maken hebben,
zoals objectagnosie (niet in staat zijn objecten te herkennen),
prosopagnosia (gezichtsblindheid), problemen kleuren te benoemen of
herkennen en dergelijke. Het zal na het bovenstaande geen verbazing
meer wekken dat men bij individuele-zenuwcelmetingen aan wakkere
taakvervullende aapjes in de temporaalkwab gevonden heeft dat er
cellen zijn, die specifiek op gezichten, kleuren of
kleurcontrasten, bepaalde objecten of objectklassen en dergelijke
reageren (Logothetis & Sheinberg, 1996). Hoewel dit alles zeer
spannend is gaan we er hier niet verder op in (zie bijvoorbeeld van
de Grind, 2000).
De `betrouwbaarheid' van het neuronaal correlaat van bewust zien
kan niet groter zijn dan van de extrastriate modules die de
informatie aanleveren. Patiënten met het zogenaamde Charles
Bonnet syndroom hebben bijvoorbeeld een dissociatie tussen de
sensorische input en de extrastriate modules. De patiënten
gaan plotseling slecht zien en krijgen visuele hallucinaties, maar
ze blijken psychiatrisch normaal te zijn. De extrastriate
cortexgebieden kunnen spontaan actief worden en zo visuele
hallucinaties veroorzaken die niet verschillen van vroegere bewuste
visuele ervaringen. Hersenscanstudies bij deze patiënten tonen
aan (ffytche et al, 1998) dat de bewuste visuele ervaringen tijdens
hallucineren overeenkomen met de rol van de extrastriate gebiedjes
die op dat moment actief zijn (kleurenfenomenen, beweging,
objectenzien, enz). Kortom, we zien hier iets terug van de zorgen
van bisschop Berkeley uit de zeventiende eeuw. Hij stelde dat we
ons bewust zijn van de interne activiteiten van het brein en niet
rechtstreeks van de buitenwereld, dus kunnen we uit de waarneming
niet weten of die buitenwereld wel echt bestaat. Gelukkig valt dat
nogal mee in dit voorbeeld. In de eerste plaats krijgen de
extrastriate cortexgebiedjes hun visuele betekenis voor een flink
deel tijdens en dankzij voorafgaande visuele ervaring met de
wereld. Zonder die opvoeding werken ze niet normaal. In de tweede
plaats kunnen deze patiënten hun hallucinaties direct van
echte waarneming onderscheiden, omdat de hallucinaties niet en de
echte waarnemingen wel worden beïnvloedt door hun hoofd- en
oogbewegingen. Bovendien zien de patiënten bij gewoon
waarnemen slecht, terwijl de hallucinaties de kwaliteit van hun
vroegere waarneming hebben. De hallucinaties zijn daarnaast vaak
bizar of zelfs lachwekkend
Laten we voor wat betreft de visuele neurowetenschap tenslotte
het verschijnsel blindzien bespreken (Engels `blindsight, Duits
`Seelenblindheit'). Blindzien is ook weer zo'n mode-onderwerp voor
denkers over bewustzijn geworden en die hebben de neiging er
gekkere conclusies aan te verbinden dan de gevonden feiten
toelaten. Blindzien kan optreden bij grote lesies in V1. De
patiënten zijn bij een lesie in de linker V1 effectief blind
in de rechter (contralaterale) gezichtshelft en vice versa. Zo'n
patiënt kan bijvoorbeeld als hij dorst heeft en er staat een
glas water in zijn blinde gezichtshelft dat glas niet zien en pakt
het dus niet op. Hij weet zeker in dat deel van hetgezichtsveld
volslagen blind te zijn en de meeste gedragstests ondersteunen dat
idee.
Als een onderzoeker de patiënt nu weet te overtuigen dat
hij bij het geluid van een belletje toch eens moet proberen aan te
geven of een dikke grote lijn in het blinde gezichtsveld
horizontaal of verticaal staat, dan blijkt de patiënt dat
vrijwel steeds goed te `raden'. Toch blijft de proefpersoon beweren
niets gezien te hebben. Hetzelfde geldt bijvoorbeeld voor visuele
bewegingen. Als de patiënt moet aangeven wat de
bewegingsrichting is `raadt' hij die vrijwel 100% goed, ondanks het
feit dat hij zeker weet nooit iets te zien. Er is dus enig `zien'
overgebleven maar dat is niet-bewust.
Wat is hiervan de neurowetenschappelijke verklaring? Zoals
Fig. 3 aangeeft is er een baan van het netvlies via de
pulvinar (PUL) naar V2 en in feite projecteert die baan ook nog op
andere extratriate gebieden, onder andere V3 en MT. Van de cellen
in MT blijkt nog ongeveer de helft redelijk normaal te functioneren
en dat blijkt dus voldoende voor niet-bewuste detectie van de
bewegingsrichting, maar niet voor bovendrempelig aansturen van het
neuronaal correlaat van bewust zien. Het verschijnsel zou daarom
opgevat kunnen worden als ondersteuning van het model van Crick en
Koch, dat het neuronaal correlaat van bewust zien niet in de
extrastriate gebiedjes op zich zetelt maar in de frontale
cortex.
Er is in dit verband nog een fysiologisch gegeven dat een
belangrijke rol speelt en dat niet in Fig. 3 is terug te
vinden. Vanuit het netvlies projecteren verschillende celklassen
naar de middenhersenen (corpus geniculatum laterale of CGL,
pulvinar of PUL, colliculus superior CS). Behalve een celklasse die
we niet zullen bespreken (W-cellen) zijn dit de parvo- of P-cellen
(de naam zegt dat ze relatief klein zijn) en de magno of M-cellen
(grote, snelle, transiënt reagerende cellen). Het lijkt er nu
op dat bij blindzienpatiënten vooral de M-banen via de
pulvinar gespaard blijven. Bovendien weten we dat de M-cellen lage
resolutie zien en bewegingszien ondersteunen, terwijl de P-cellen
vooral domineren bij hoge resolutie vormzien, kleurenzien en
dergelijke. Dit klopt met de bevinding dat de blindzieners in hun
blinde gezichtsveld lage resolutie hebben, dus alleen de
oriëntatie van zeer grove lijnen goed kunnen raden en dat
kleurcontrast ze niet helpt bij beter onderscheiden van voorgrond
en achtergrond. Een bepaalde patiënt van L.Weiskrantz in
Oxford kon beweging zelfs bewust waarnemen in zijn blinde
gezichtsveld als het bewegende object of de textuur maar van heel
hoog contrast was. Bij geleidelijk verlagen van het contrast
verdween de bewuste waarneming, maar kon de patiënt nog lange
tijd vrijwel perfect `raden' in welke richting de beweging ging. Er
was dus een graduele overgang van bewust naar niet-bewust zien.
Blindzien heeft men ook bij aapjes die V1 missen kunnen
aantonen. Ze werden eerst getraind om met een hendeltje aan te
geven in welke richting een beweging plaats vond en op twee knoppen
te scoren of er een visuele stimulus optrad na een belletje of
niet. Na verwijdering van V1 rechts hadden ze dus in principe links
van het midden alleen blindzien. Als de onderzoekers de aapjes dan
lieten aangeven in welke richting een beweging verliep dan scoorden
de aapjes vrijwel 100%, ook in hun blinde veld, terwijl ze bij de
stimuli in hun blinde veld op de knoppen tegelijk aangaven dat er
geen prikkel was geweest! De verschijnselen zijn in principe bij
aap en mens gelijk en dit type experiment biedt de mogelijkheid dit
ook bij andere zoogdieren (bijvoorbeeld katten) na te gaan.
Deze bevindingen zijn in de filosofische literatuur nogal wat
opgerekt. Daniel Dennett stelt zich bijvoorbeeld in een
gedachtenexperiment voor dat we een blindzienpatiënt zouden
kunnen trainen te raden wanneer hij moet raden. We zouden die
persoon, aldus Dennett, dan niet van een normaal-ziende kunnen
onderscheiden en we zouden dus een persoon hebben die kan zien
zonder bewuste perceptie, een zombie. Dit is een aardig voorbeeld
van een doorgeschoten gedachtenexperiment. Immers, de blindziener
is helemaal geen normaal-ziende verminderd met bewust beleven. In
het blinde gezichtsveld is niet alleen de resolutie enorm laag,
maar de patiënt detecteert er op zijn best beweging en de
oriëntatie van zeer grote lijnen/balken. Tot op heden is er
nog nooit echt vormzien gerapporteerd in blindzienpatiënten
(Young & de Haan, 1993) en ook kleurenzien is bij verreweg de
meeste blindzienpatiënten afwezig en bij anderen rudimentair.
Voor een gedetailleerdere behandeling van het probleem van
oversimplificatie van het verschijnsel blindzien ten behoeve van
een vlotte theoretische generalisatie verwijzen we naar Tapp
(1997).
Het verschijnsel van blindzien kan mogelijk het beste worden
vergeleken met wat we allemaal weleens beleven bij sterke mist of
zeer lage lichtsterkte, namelijk dat we menen iets te zien, kunnen
raden wat het is, maar dat raden niet echt als bewuste visuele
herkenning ervaren. Ook in formele psychofysische experimenten
vindt men vaak een klein parameterbereik, zeg van het contrast,
waar de proefpersoon nog consistent vrijwel 90-100% goed raadt,
maar subjectief van mening is dat hij/zij volledig willekeurig
antwoordt. Dit lijkt sterk op het bovenbeschreven verschijnsel bij
de patiënt van Weiskrantz. Er is een gebied van niet-bewust
zien waar de stimulusattributen boven kansniveau worden geraden,
ook al menen we ze niet echt te `zien'.
Uit het bovenstaande volgt naar onze mening duidelijk dat het
empirisch neurowetenschappelijk onderzoek aan bewuste
verschijnselen goed op gang is gekomen en al bijzonder interessante
gegevens heeft opgeleverd. Het is gebleken dat men moet oppassen
voor het trekken van te ver gaande conclusies uit losse
gedachtenexperimenten gebaseerd op een globale samenvatting van de
bevindingen. De meeste experimenten zijn complex en dienen met veel
gevoel voor detail te worden geïnterpreteerd, liefst door
denkers met ervaring in neurowetenschappelijk onderzoek. Een kop
als `zinloos bewustzijn' boven een artikel in een
wetenschapsbijlage van een landelijke krant (NRC, 11 september
1999) en de vele vrije speculaties in dat artikel kunnen dan
hopelijk in de toekomst worden vermeden of vervangen door meer
gefundeerde samenvattingen van resultaten van feitelijk
onderzoek.
Gegeneraliseerd blindzien levert geen zombie op maar gewoon een
blinde. Bewustzijn is niet zinloos, maar noodzakelijk voor de
overleving. Vrije wil is geen illusie maar het gevoel dat hoort bij
de activiteit van actie-initiërende netwerken in het
zenuwstelsel en dat gevoel komt niet achteraf als de niet-bewuste
beslissingen over detailbewegingen al zijn genomen, maar ervoor als
de beslissing om aan het spel mee te doen valt. Het Cartesiaanse
theater, de vrije wil en het werkgeheugen zijn metaforen die
wellicht beter gezamenlijk door de metafoor `bedrijfssysteem' of
`directiekamer' vervangen kunnen worden. Ze kunnen dan gemakkelijk
van hun metafysische lading worden ontdaan en onderdeel worden van
wetenschappelijke theorieën die toetsbare voorspellingen
leveren. Het neuronaal correlaat van bewustzijn bestuurt namelijk
volgens de moderne inzichten lang niet alles wat er in het
zenuwstelsel gebeurt en houdt evenmin alles in de gaten. Zodoende
kan het niet worden vergeleken met de niet-materiële geest van
Descartes en evenmin als een homunculus (mensje in de mens) worden
opgevat.
Met dit alles is de oplossing van wat wel `het moeilijke
probleem' wordt genoemd (waarom en hoe geeft hersenactiviteit
eigenlijk aanleiding tot mentale fenomenen?) niet enorm veel
naderbij gebracht. Toch moeten we ons daardoor niet laten
ontmoedigen. Een bepaald type problemen heeft er een handje van om
ongemerkt van het toneel te verdwijnen. Het Kracht-Stof Probleem is
hier een goed voorbeeld van. Heel negentiende-eeuws geleerd Europa
zat er mee in zijn maag, maar tegenwoordig maakt niemand zich er
nog druk over. Er is geen reden om te veronderstellen dat het met
het tegenwoordige `geest-lichaam probleem' anders zit. Op een goede
dag zullen er wellicht zoveel feiten en interessante theorieën
voorhanden zijn dat niemand zich er nog om bekommert.
Wellicht is het aardig af te sluiten met enige speculaties van
Ramachandran en Hirstein (1997). Zij stellen dat het moeilijke
probleem vooral moeilijk is of lijkt, omdat we een aantal
vertaalslagen naar andere media doorlopen. Persoon A vertelt over
zijn roodervaring en persoon B probeert zich daarin `in te leven'.
We gaan dan van activiteiten in bijvoorbeeld gebied V4 van persoon
A naar de taalcortex, naar spraakspieren, luchttrillingen,
gehoorsysteem van B, decoderen van de neuronale activiteit van het
gehoorsysteem in de auditieve cortex en een poging van B om dat op
te vatten als activiteit in zijn eigen V4. Volgens Ramachandran
& Hirstein praten we dus `slechts' over onmogelijke
vertalingen. Als we V4 van persoon A neuron voor neuron op V4 van
persoon B zouden kunnen aansluiten zou B wellicht direct bij de
roodervaring van A uitroepen `oh, wat mooi rood'. Als zij gelijk
zouden hebben zou dat een reden temeer zijn om voorlopig maar
driftig onderzoek aan de neuronale mechanismen van bewustzijn te
doen en niet te veel tijd te besteden aan gepraat over het
onbespreekbare.
- Baars BJ (1998). Metaphors of consciousness and attention in
the brain. Trends in Neuroscience 21 (2), 58-62
- Britten KH & van Wezel RJA (1998). Electrical
microstimulation of cortical area MST biases heading perception in
monkeys. Nature Neuroscience Vol. 1, no. 1, 59-63. (See
also the editorial on this work in the same issue by B.Cumming, A
sense of direction: going with the flow, pp. 6-8).
- Clarke E & Dewhurst K (1972). An Illustrated History of
Brain Function. University of California Press, Berkeley and Los
Angeles
- Clarke E & O'Malley CD (1968). The Human Brain and Spinal
Cord: A Historical Study Illustrated by Writings from Antiquity to
the Twentieth Century. University of California Press, Berkeley and
Los Angeles.
- Courtney SM, Petit L, Maisog JM, Ungerleider LG, Haxby JV
(1998). An area specialised for spatial working memory in human
frontal cortex. Science 279, 1347-1351.
- Crick F & Koch C (1990). Towards a neurobiological theory
of consciousness. Semin. Neuroscience 2, 263-275.
- Crick F & Koch C (1995). Are we aware of neural activity in
primary visual cortex? Nature 375, 121-123 & 294-295.
- Crick F & Koch C (1998). Feature article: Consciousness and
neuroscience. Cerebral Cortex 8, 97-107.
- Dennett DC, Kinsbourne MJ (1992). Time and the observer: the
where and when of consciousness in the brain. Behav. Brain Sci 15,
183-247.
- Dijksterhuis EJ (1950). De mechanisering van het wereldbeeld.
Meulenhoff, Amsterdam.
- Finger S (1994). Origins of Neuroscience. Oxford University
Press, New York and Oxford.
- ffytche DH, Howard RJ, Brammer MJ, Woodruff P & Williams S
(1998). The anatomy of conscious vision: an fMRI study of visual
hallucinations. Nature Neuroscience 1 (8), 738-742.
- Goldman-Rakic PS (1992). Working memory and the mind.
Scientific American 267 (3), 73-79
- Gomes G (1998). The timing of conscious experience: A critical
review and reinterpretation of Libet's research. Consciousness
& Cognition 7, 559-595.
- Grind WA van de (1997). Natuurlijke Intelligentie. Uitgeverij
Nieuwezijds, Amsterdam.
- Grind WA van de (2000). Bewust zien en zijn (deel 1).
Neuropraxis 4 (1), 9-15.
- Keller I & Heckhausen H (1990). Readiness potentials
preceding spontaneous motor acts: voluntary versus involuntary
control. Electroencephalography and clinical Neurophysiology 76,
351-361
- Kim J (1996). Philosophy of Mind. WestView Press, Boulder,
Colorado.
- Kornhuber HH & Deecke L (1965).
Hirnpotentialänderungen bei Willkürbewegungen und
passiven Bewegungen des Menschen: Bereitschaftspotential und
reafferente Potential. Pflügers Arch ges Physiol 284,
1-17.
- Lecours AR & Joanette Y (1980). Linguistic and other
aspects of paroxysmal aphasia. Brain Lang. 10, 1-23.
- Libet B (1985). Unconscious cerebral initiative and the role of
conscious will in voluntary action. Behavioral and Brain Sciences 8
(4), 529-566.
- Libet B (1993). The neural time factor in conscious and
unconscious events. In: Experimental and theoretical studies of
consciousness. Ciba Foundation Symposium 174, 123-146.
- Logothetis NK & Sheinberg DL (1996). Visual object
recognition. Annual Review of Neuroscience 19, 577-621.
- Lokhorst GJC (1986). Brein en bewustzijn: de geest-lichaam
theorieën van moderne hersenonderzoekers. Uitgeverij Eburon,
Delft.
- Lokhorst GJC (1996). Counting the minds of split-brain
patients. Logique et Analyse 39, 316-324.
- Lokhorst GJC & Kaitaro TT (te verschijnen). The originality
of Descartes' theory about the pineal gland. Journal of the History
of the Neurosciences, te verschijnen.
- Milner AD & Rugg MD (1992). The neuropsychology of
consciousness. Academic Press, New York.
- Parker AJ & Newsome WT (1998). Sense and the single neuron:
Probing the physiology of perception. Annual Review of Neuroscience
21, 227-277.
- Ramachandran VS & Hirstein W (1997). Three laws of qualia:
What neurology tells us about the biological functions of
consciousness. Journal of Consciousness Studies 4 (5-6),
429-457.
- Rasmussen JL, Rajecki DW & Craft HD HD (1993). Humans'
perceptions of animal mentality--ascriptions of thinking. J Comp
Psychol 107: 3, 283-290.
- Salzman CD, Britten KH & Newsome WT (1990). Cortical
microstimulation influences perceptual judgments of motion
direction. Nature 346, 174-177.
- Searle JR (1984). Minds, brains and science. Harvard University
Press, Cambridge Mass.
- Searle JR (1992). The rediscovery of the mind. MIT Press,
Cambridge Mass
- Searle JR (1998). How to study consciousness scientifically.
Phil.
Trans. R. Soc. Lond. B 353, 1935-1942.
- Tapp TD (1997). Blindsight in hindsight. Consciousness &
Cognition 6, 67-74
- Verbeke G (1945). L'évolution de la doctrine du pneuma
du stoïcisme à S. Augustin. Desclée de Brouwer,
Paris, & Éditions de l'Institut Supérieur de
Philosophie, Louvain.
- Went FW (1968) The size of man. American Scientist 56 (4),
400-413.
- Young A & de Haan EH (1993). Impairments in visual
awareness. In M. Davies & G.W. Humphreys (Eds), Consciousness.
Blackwell, Oxford.
- Zeki S & Bartels A (1998). The autonomy of the visual
systems and the modularity of conscious vision. Phil. Trans. R.
Soc. Lond. B 353 (1377), 1911-1914.
Previous | Up |
Next
gjclokhorst@gmail.com || July 17, 2015 || HTML 4.01 Strict